THE RESEARCHERS sat in a Land Rover just outside southeastern Kenya’s Tsavo East National Park, waiting. Hours ticked by. Nearby, a fresh zebra carcass lay only half eaten.
But the lions that live in this largely unstudied section of the country, unlike wildlife more accustomed to human observers, were wary. Hiding in the thick thornbush that blankets the area, they refused to return to their kill.
It grew darker. Finally, after four hours, one of the lionesses was apparently fed up. With a terrifying, blustering snarl, she suddenly charged. The scientists snapped to attention; they knew of the Tsavo lions’ reputation as oversized brutes. In 1898, two of these predators reportedly attacked, killed and ate 135 men building a railroad across Kenya.
In the end, the lion pulled up just short of the Land Rover. Then the pride’s leader, a male called Cassius, did something remarkable. Accompanying three cubs, he escorted them to the zebra and kept a watchful eye as the youngsters ate. "That is totally unknown in the Serengeti," says biologist Bruce Patterson, lead researcher on a project to study the lions.
Why Cassius took it on himself to guard the cubs—instead of leaving the task to females in the pride—is anyone’s guess. But it’s not the only mystery surrounding Tsavo lions. Are they, for example, a separate species from Panthera leo living elsewhere on the continent? Why do so many male Tsavo lions lack manes? Are they bigger and more aggressive than other lions? And are the predators really man-eaters? These are the kinds of questions Patterson and his colleagues are attempting to answer about a group of animals that has fascinated observers since their notorious rampage a century ago.
Because they’re far less studied than lions on the Serengeti plains of Kenya and Tanzania, much of what’s known about Tsavo lions is based on speculation, not science. "I’d like to set the record straight about what these lions are really about," says Patterson, curator of mammals at Chicago’s Field Museum and author of The Lions of Tsavo: Exploring the Legacy of Africa’s Notorious Man-Eaters.
Much of the relatively recent interest in Tsavo lions can be traced directly to a display in a back corner of Patterson’s museum. One summer afternoon, a tour group paused outside the glass-and-wood case housing two adult Tsavo lion specimens. Sleek and lean, they look unremarkable except in one way: The lions have no manes. Compared to a typical zoo lion, they look virtually bald. The guide then told the story of the predators’ man-eating spree, a tale that’s been the subject of three books and two movies, including a 1996 feature film, The Ghost and the Darkness.
"Tsavo," the guide began, "means ‘place of slaughter.’" Between March and December of 1898, he said, the two lions in the display case terrorized Indian workers brought by the British to build a rail line from Lake Victoria to the Indian Ocean. By the end of their rampage, the predators had stalked, seized and eaten 135 laborers. Many of the victims were dragged from tents during the night and devoured within earshot of camp. "They ate people like Twinkies," the guide said. "And they killed for pleasure."
Construction at Tsavo was halted until December 1898, when John Patterson, the project’s lead British engineer, finally shot and killed both lions. Patterson (no relation to Bruce Patterson) went on to tour the world, telling his story. Eventually, he sold the lions’ skins to the Field Museum, where they were stuffed and mounted. Viewed by hundreds of thousands of visitors each year, they remain one of the museum’s most popular attractions.
The story of the Tsavo lions remained unexamined for nearly 100 years, when Field Museum scientists began to look into it. In 2000, researcher Tom Gnoske discovered that the railroad office in London had records of only 28 deaths of foreign workers. Though it’s possible that the lions killed many more Africans—whose deaths were not recorded—there’s no evidence that the total reached John Patterson’s claim of 135. "It’s part of the myth," Gnoske says.
Gnoske and his colleagues also explored whether the lions might have had an inherent taste for human flesh. On the contrary, they found that the predators may have been forced to attack people when an outbreak of rinderpest between 1891 and 1893 decimated buffalo herds, their usual prey. Scavenging along the Tsavo caravan route, the lions would have found bodies of dead or dying slaves abandoned by ruthless slave traders. Other bodies were left more or less exposed by local burial practices. Lions are quick learners, and it would not have been a big leap to then turn to living humans.
Another possibility is that a nagging toothache could have led at least one of the predators to prey on people. A radiograph image of one specimen’s skull reveals a severe root tip abscess. "Any sort of pressure on the area would have been extremely painful,’’ Patterson says, making it difficult for the lion to deliver a "killing bite" to either the nape or throat of prey. Humans, no doubt, presented a less taxing meal.
Though no people-eating rampages have been reported since the late 1800s, conflicts between man and beast continue. Records from the Kenya Wildlife Service show 112 attacks on cattle, sheep or goats between 1994 and 1998 in Tsavo East, one of two parks within a larger protected area. During that period six humans were also attacked—and two were killed. The problem is worse outside the parks. On the 96,000-acre Taita Ranch and another nearby ranch, where Patterson currently conducts research, lions attacked livestock nearly 300 times between 1996 and 1999. Across the entire 1.5 million-acre zone surrounding the protected area, lions attack livestock as often as once per day, estimates Steve Turner, a trustee for Kasigau Conservation Trust, which is working to create a wildlife corridor between parks.
And there are still reports of human attacks. In June 2002, for example, lions killed a herdsman along a highway to the coastal city of Mombasa. Usually, however, such assaults are more fatal to the predators than to people. So-called "problem" lions are routinely captured or shot. One animal control officer from Tsavo reported shooting 222 lions across Kenya over 12 years. "Conflict with humans is the number one mortality factor for African lions," says Roland Kays, mammal curator at the New York State Museum and a partner in Patterson’s research.
Many locals still believe that old and sickly lions, possibly with tooth problems, are responsible for most human attacks today. While that might have been the case in the 1898, Field Museum researchers have found that attacking lions these days are typically under five years old and healthy.
Another hypothesis Patterson’s team is exploring is whether Tsavo lions have elevated levels of testosterone. More hormones might lead males to vigorously defend larger territories, leaving less room for youngsters. It could also lead to a condition similar to male-pattern baldness in people, when testosterone receptors on hair follicles are overloaded and cause hair loss, contributing to the absence of manes on the lions. Patterson and colleagues are now collecting hair follicles for analysis by a British endocrinologist.
Testosterone might also explain another phenomenon: single males living with larger prides. In a survey of the lion population of Tsavo East, Patterson and Kays found that average pride size was 7.4 lions compared to 6.4 in the Serengeti. And Tsavo prides typically had just one male, while those in other areas tended to have two or more. How would individual males fend off large groups of wandering males? "It’s still a mystery to us," Patterson says. "But if they had more testosterone they might be aggressive enough to hold onto a pride."
That theory is at odds, though, with research conducted by University of Minnesota biologist Craig Packer, a leading expert on Serengeti lions, and his former student Peyton West. These scientists note that castrated lions, or lions with injuries to their genital regions, lose their manes. Other lions without manes are typically younger or sickly and almost always subordinate to maned males.
Regardless of hormone levels, environmental factors likely play a greater role. Gnoske and biologist Julian Kerbis Peterhans of the Field Museum and Roosevelt University in Chicago compiled data on lions from 300 different locations. They found that the higher the altitude and cooler the temperatures, the bigger the manes. "There are other factors involved but we think the most important one is this temperature business," Kerbis Peterhans says.
Another variable is water, or its severe shortage over long periods of time. In a part of Tsavo East where maneless lions are common, annual rainfall is just 12 inches. On Taita Ranch, however, there is significantly more rain. There lions like Cassius feature what Patterson calls a modest mane: a mohawklike growth on the head, hair on the neck and chest, but bare shoulders. In parts of the Serengeti, where all males sport full manes, annual rainfall can be nearly four times as much as it is in Tsavo.
Unable to cool down during the dry season—which can last four to six months in Tsavo—a maned lion would be a less efficient hunter, having to skip the hottest parts of the day and remain close to small areas near permanent water sources. That challenge could trigger a hormonal response limiting mane growth. There’s no doubt that a mane makes a lion hot: Thermal images taken by West in the Serengeti show male lions were hotter than females; in Tsavo, maneless males were not hotter.
But West isn’t sold on the idea that manelessness is that prevalent in Tsavo. Because of the harsh climate and thornbush terrain, lions might develop manes later in life, she says, or might continually lose them and grow them back. And they might simply be less regal: "It may be the biggest mane that any Tsavo lion could grow won’t be as big as any mane a Serengeti lion could grow."
While the mane question remains unsolved, Patterson believes he has dispelled other popular ideas about the lions: that they are bigger or possibly a separate species or subspecies of African lion. Charting the skull size of 18 Tsavo lions, he found that they ranged from more than 11 inches to nearly 15 inches—well within the range of lions elsewhere on the continent. And an analysis of the lions’ DNA revealed that their genetic make-up is virtually identical to that of Panthera leo throughout Africa.
Whatever else his research unveils, Patterson hopes his project will help ensure that the lions remain in Tsavo for decades to come. Through Earthwatch Institute, a Massachusetts-based conservation group, volunteers pay fees to visit Tsavo and help Patterson conduct his fieldwork. A portion of their fees goes directly to Kenyan landowners who cut back on grazing, leaving more room for wildlife, including some 30 lions and 200 elephants. Without the volunteers’ contributions, totaling about $35,000 a year, Turner believes the land would be home to "10,000 cows and probably nothing else."
Still, there are limits to this kind of conservation. When a terrorism scare hit Kenya in the summer of 2003, Earthwatch canceled several trips and overall tourism to the country dropped. But Patterson is undeterred. "The battle to save this region’s wildlife," he says, "is going to be won or lost in Tsavo."
Chicago writer Dave Newbart wrote about China’s black-necked cranes for the January/February 2001 International Wildlife.
Wednesday, November 21, 2007
Tsavo
Back in 1898, the British decided to build a railroad in East Africa. This railroad would stretch from Mombasa on the coast of modern-day Kenya to Lake Victoria, and later into Uganda. This railroad, called the Uganda Railroad was also referred to as 'The Lunatic Line'. It was said by it's opponents to go 'from nowhere to utterly nowhere'.March 1898 the British started building a railway bridge over the Tsavo (SAH-vo) River in East Africa. Over the next nine months, two large male lions killed and ate nearly 140 railway workers. Crews tried to scare off the lions and built campfires and thorn fences for protection, but to no avail. Hundreds of workers fled Tsavo, halting construction on the bridge.Before work could resume, chief engineer Lt. Col. John Henry Patterson (1865-1947) had to eliminate the lions and their threat. After many near misses, he finally shot the first lion on December 9, 1898, and three weeks later brought down the second. The first lion killed measured nine feet, eight inches (3 m) from nose to tip of tail. It took eight men to carry the carcass back to camp. The construction crew returned and completed the bridge in February 1899.
Nevertheless, this railroad had some legitimate purposes to exist. In those days, the only route into the interior of the African continent was on foot. There was much in the way of agricultural goods and other wares that could not easily reach market due to the lack of transportation. Missionaries had considerable trouble reaching the rich 'fields' in central Africa. Finally, there was was the nagging slave trade problem. It was hoped that good transportation would encourage people living in the interior of the continent that there were better ways of making a living than capturing slaves.After completing the railroad, Patterson became chief game warden in Kenya and later served with the British Army in World War I. He published four books and lectured widely on his adventures. After speaking at The Field Museum in 1924, Patterson sold the museum the lion skins and skulls for the then-sizeable sum of $5,000.
The skins arrived in less-than-perfect condition--old and dry, they had been cut down into rugs. (In real life the lions were even larger than they appear as taxidermy mounts.) The skins were also blemished by gunshot wounds and thorn scratches. Museum taxidermist Julius Friesser did an extraordinary job creating the life-like mounts you can see at The Field Museum.
The construction of this railroad remains one of the great engineering feats of the late 1800's. It's 580 miles of track had to cross the great rift valley, several rivers, and some of the most inhospitable territory you could imagine. Construction started in 1896, and reached what is today Nairobi in 1899. It finally reached Kismu on Lake Victoria in 1901. It took 27 more years for the railroad to actually be extended to Kampala, Uganda.
It was not many days after Colonel Patterson arrived that reports started coming in about workers disappearing. Although it was told him that lions were responsible, Patterson at first didn't believe it. When he finally investigated, it was quickly (and gruesomely!) discovered that not one but two lions were responsible for killing the workers. Early attempts to shoot the lions were unsuccessful. The lions seemed to be able to predict what Patterson would do next. And, with camps scattered up and down the railway for 30 miles, the lions could strike in a different camp each night
It was not many days after Colonel Patterson arrived that reports started coming in about workers disappearing. Although it was told him that lions were responsible, Patterson at first didn't believe it. When he finally investigated, it was quickly (and gruesomely!) discovered that not one but two lions were responsible for killing the workers. Early attempts to shoot the lions were unsuccessful. The lions seemed to be able to predict what Patterson would do next. And, with camps scattered up and down the railway for 30 miles, the lions could strike in a different camp each night.
It was not many days after Colonel Patterson arrived that reports started coming in about workers disappearing. Although it was told him that lions were responsible, Patterson at first didn't believe it. When he finally investigated, it was quickly (and gruesomely!) discovered that not one but two lions were responsible for killing the workers. Early attempts to shoot the lions were unsuccessful. The lions seemed to be able to predict what Patterson would do next. And, with camps scattered up and down the railway for 30 miles, the lions could strike in a different camp each nightThe workers soon constructed thorn bomas (A boma is a thorn fence erected around a dwelling or corral designed to keep predators such as lions out.) around their camps, and kept fires burning at night to try to scare off the lions. But it didn't work. The lions literally ignored the thorns, and would drag themselves and their meals right through them. This caused no small concern among the workers.Still, there were a couple of lucky escapes. One night, a lion attacked a man riding a donkey. The donkey was knocked over and the man knocked off. The lion moved in for the kill, and somehow got his claws hooked on a rope tied to some oil cans that had been around the donkey's neck. The lion couldn't immediately figure out how to unhook the rope, and the oil cans were making a terrible racket. The noise frightened the lion so much that it ran off back into the bush, dragging the oil cans with it. The rider escaped to the safety of a tree and stayed there the rest of the night.
Another time, one of the lions broke into a tent, and was intent on carrying off the occupant who was sleeping on a mattress. Instead, somehow, the lion got hold of the mattress and pulled it out from under the man. Soon realizing it's mistake, the lion dropped the mattress and ran off. Another time, one of the lions jumped onto a tent containing 14 Indian 'Coolies'. The lion broke through the tent, clawing up one man's shoulder in the process. Somehow in the ensuing confusion, the lion grabbed a sack of rice and made off with that instead. The lion 'threw it down in disgust' a short distance away and beat a hasty retreat.
At first, the effect of the lions grisly habits on the workers wasn't too great, because the camps were spread out over a large area. But, a large area. But, as the railhead pushed Northwestward, only a few hundred workers were left at Tsavo to build the bridge. The lions now directed their efforts on this one camp, and this is when the morale at Tsavo really began to drop! One night, one of the lions attacked the hospital tent. At first, he was scared away when the doctor's assistant knocked over a cabinet of supplies in fright. But, the lion tried again. Breaking through the tent, he seized one of the patients, and injured 2 of the other patients. It was decided after that to move the hospital tent. The very next night, a lion attacked the new hospital tent! Many of it's occupants got to witness the lion seize, kill and drag through the thorn boma the hospital's water-carrier. The next day, (as was usually the case for these lions) all that was left of the water-carrier was his head, a few of the larger bones, and part of his hand.The hospital-tent was moved again, and an even thicker boma was erected around it. A supply wagon was parked at the previous hospital tent location, and some cattle were tied up inside the boma as bait. Colonel Patterson and the Doctor stayed up all night, hoping to get a shot at the lions. Their vigilance was rewarded, as one lions jumped into the boma. Patterson and the doctor figured that the lion had silently dispatched one of the cattle and was trying to find a place to get out of the boma. Unbeknownst to them, the front entrance to the boma was not properly secured, and the lion exited by this route. It then proceeded to stalk Patterson and the Doctor. Luckily for them, they figured this out in time. The lion attacked, and Patterson got off a shot. This scared off the lion. Colonel Patterson writes much later in the Field Museum account of that incident that he believes that he shot one of the canine teeth out of the mouth of that lion. If this was indeed the case, this lion was the one now known as maneater #1. The date was April 23rd.
After the incident at the old hospital site, the lions left the area for a few months, and later accounts report people were disappearing at other camps some miles away. During this time, a considerable amount of effort was spent building an elaborate trap using a boxcar, to be ready should the lions return. The boxcar was divided into two compartments by strong bars. A trip wire was arranged to drop a gate made of railroad rails over the entrance to one of the compartments. (For lack of a suitable drill, Colonel Patterson 'drilled' holes in the rail sections with his .303 rifle!) There was a small, secure entrance into the other compartment. Thorns were piled all around the boxcar. For the first few nights, Colonel Patterson sat inside the secure compartment, acting as human bait. Should a lion sense his presence inside the boxcar, it would enter and trip on the trip wire. The gate would fall and trap the lion in the first compartment. The Colonel could dispatch the lion at his leisure from behind the strong bars. After a few days, he got some other people to take turns manning this trap.
The lions had been gone for so long that the workers started to let down their guard a little. This didn't last long. One night, some of the Coolies had decided to sleep outside their tent, but inside the boma. This would not be their lucky night. One of the lions jumped into the boma. Although stones and firebrands were hurled at the lion, it selected a victim, and dragged it out of the boma through the thorns! Outside the boma, the other lion joined the first one, and they enjoyed their meal not 30 yards away. Even though shots were fired at the lions, none of them made their mark, and the lions went right on eating.Soon, a sort of routine would settle in. The lions would be heard roaring in the vicinity of one of the camps (They were again spread out up and down the rail line.). Suddenly, they would stop roaring. Word would go from camp to camp, 'Beware brothers, the devil is coming!'. There would be agonizing shrieks from somewhere in the camp, and one less person at roll call the next morning.Every night, Colonel Patterson and others would stay up, hoping to get a shot at one of the lions. They never did.
The lions were getting bolder and bolder. Some nights, they would each take a victim, so they wouldn't have to share. They could go undetected right through the thorn fences. One night, a bunch of Coolies escaped the lions to the safety of a tree. They so heavily loaded the tree that it collapsed, throwing them to the ground very close to the lions. But, the lions didn't care. They had already caught a victim and were too busy feasting on him.
One night, the lions caught a victim and carried it close to Colonel Patterson's camp to devour. He vividly remembers the sound of bones being crunched and contented 'purring'. It took days to get these sounds out of his head. (Lions actually can't purr like smaller felines. They have a rough 'pseudo purr' they don't often use.) Finally, on December 1st, the workers approached Colonel Patterson and told him they would no longer be 'food for lions or devils'. As soon as this ultimatum was delivered, the workers went out and threw themselves down on the tracks in front of the next train. They then climbed on every available seat, and left Tsavo. The entire railway project ground to a halt. Only a small number of workers were brave enough to stay behind. These workers took up residence in trees, atop water in trees, atop water tanks, or in underground pits covered with logs.
On December 3rd, the Superintendent of Police arrived with twenty men to help hunt down the lions. It was on that night that one of the lions finally entered the rail-car trap. There were three armed men in the secure part of the trap. After the gate had fallen, the lion began to thrash at the bars, scaring the men silly. Finally, they got their courage up and began firing. Even though they could have touched the body of the lion with the muzzles of their rifles, nobody succeeded in getting a shot into it. Instead, one of the shots broke the chain holding the gate shut. The lion escaped with very minor injuries. The superintendent of Police, his men, Patterson and others tried to track the lions. Although one person got a brief glimpse, they again escaped! They tried again for two more days to track the lions. No success. The Superintendent and his men could not afford to stay any longer. Patterson was again left alone with the lions.
Patterson then rounded up his remaining Coolies, and instructed them to bring along things to make noise with. They surrounded the thicket. Patterson then stood at an animal trail coming out of the thicket. On his signal, the Coolies advanced, making all sorts of racket. It was then that Colonel Patterson actually saw one of the maneaters for the first time- a huge maneless male. He lifted his weapon to fire. Click! The twin-barreled rifle misfired! Intimidated by the noise, the lion jumped past Patterson and started to run off. He finally remembered he had another shot in his rifle and fired. He managed to hit the lion in the back, but it got away.
Dismayed, Colonel Patterson went back and looked at the donkey. The lion had just began it's meal on the hindquarters. (Lions like to start eating on the hindquarters because there is a lot of meat there without a lot of bones.) A nice donkey like this would be too good of a meal to abandon. The lion might return to finish his meal. Then, Patterson had an inspiration. Taking 4 poles, and a plank of wood, he constructed a platform in close to the donkey carcass. This platform, called a machan, would serve as an artifical 'tree' from which to hunt the lion. (There were no nearby trees to sit in.) He lashed the donkey carcass to a nearby tree stump with wire, so it couldn't be dragged off. Normally, Patterson would stand vigil with his gun-bearer (Colonel Patterson was an experienced big game hunter.), but tonight, he was sick. So, he began that night's vigil alone. It wasn't long before Patterson heard a twig snap and a sigh of hunger. The lion had indeed returned! But, as he watched, he determined the lion had detected him. Now, the lion was stalking Colonel Patterson! For two hours, the lion circled the rickety platform. All the lion would have to do is knock out a pole, or jump at the platform to dislodge Patterson. Instant easy meal! The whole situation made Patterson's flesh crawl.
Suddenly, something hit Patterson on the head! It turned out to be an owl that had confused him for a tree branch. He quickly recovered his wits. Below, the lion growled, and moved in for the kill! Very carefully, Patterson raised his rifle and fired. There was an angry growl, and the lion jumped around all over. He jumped into the bushes. Patterson fired away into bushes. The growls continued, but were growing weaker. They finally ceased altogether. One of the maneaters were finally dead!
Word quickly spread to the camps, and a wild celebration ensued. The next morning, the body of the lion was recovered. He was a maneless . He was a maneless male, 9 feet, 8 inches long from tip of nose to tip of tail. The lion had taken two shots-- one in the shoulder penetrating the heart (Probably the first shot), and another in one of the hind legs. It took 8 men to carry it to camp, and soon it was in the skinning shed The next morning, Colonel Patterson, his gun-bearer and a native tracker set off after the lion. The bloody trail was easy to follow. About a quarter mile away, they found him. He was hiding in the grass, glaring at the men with bared teeth.
Patterson aimed carefully, and took a shot. The lion then charged him with all the muster it had left. Another shot knocked him over, but he got up and continued his charge. A third shot had no effect. He reached for another rifle, only to discover his gun-bearer had abandoned him to the safety of a tree. Patterson had no choice but to follow suit. If one of the shots hadn't broken a hind leg, Patterson would have never made it. Once in the tree, the Colonel grabbed a carbine from his gun-bearer and shot the lion once more. The lion collapsed.
Rather foolishly, Patterson quickly climbed down. To his complete surprise, the lion charged again! A shot in the chest and another in the head (Look at the skull on the pictures page) finally did the brute in. Even so, the lion bit savagely at a branch until the last bit of life drained out of him.
Colonel Patterson had all he could do to prevent the dead lion from being torn apart by the workers. He had it carried to his nearby boma for examination. This lion (also a maneless male) had taken at least six shots. There was also a slug buried not far into the flesh of the back. This was the slug from the shot fired 10 days before. In any case, this lion measured 9 feet, 6 inches from tip of nose to tip of tail. Soon, this lion took it's turn in the skinning shed.
Nevertheless, this railroad had some legitimate purposes to exist. In those days, the only route into the interior of the African continent was on foot. There was much in the way of agricultural goods and other wares that could not easily reach market due to the lack of transportation. Missionaries had considerable trouble reaching the rich 'fields' in central Africa. Finally, there was was the nagging slave trade problem. It was hoped that good transportation would encourage people living in the interior of the continent that there were better ways of making a living than capturing slaves.After completing the railroad, Patterson became chief game warden in Kenya and later served with the British Army in World War I. He published four books and lectured widely on his adventures. After speaking at The Field Museum in 1924, Patterson sold the museum the lion skins and skulls for the then-sizeable sum of $5,000.
The skins arrived in less-than-perfect condition--old and dry, they had been cut down into rugs. (In real life the lions were even larger than they appear as taxidermy mounts.) The skins were also blemished by gunshot wounds and thorn scratches. Museum taxidermist Julius Friesser did an extraordinary job creating the life-like mounts you can see at The Field Museum.
The construction of this railroad remains one of the great engineering feats of the late 1800's. It's 580 miles of track had to cross the great rift valley, several rivers, and some of the most inhospitable territory you could imagine. Construction started in 1896, and reached what is today Nairobi in 1899. It finally reached Kismu on Lake Victoria in 1901. It took 27 more years for the railroad to actually be extended to Kampala, Uganda.
It was not many days after Colonel Patterson arrived that reports started coming in about workers disappearing. Although it was told him that lions were responsible, Patterson at first didn't believe it. When he finally investigated, it was quickly (and gruesomely!) discovered that not one but two lions were responsible for killing the workers. Early attempts to shoot the lions were unsuccessful. The lions seemed to be able to predict what Patterson would do next. And, with camps scattered up and down the railway for 30 miles, the lions could strike in a different camp each night
It was not many days after Colonel Patterson arrived that reports started coming in about workers disappearing. Although it was told him that lions were responsible, Patterson at first didn't believe it. When he finally investigated, it was quickly (and gruesomely!) discovered that not one but two lions were responsible for killing the workers. Early attempts to shoot the lions were unsuccessful. The lions seemed to be able to predict what Patterson would do next. And, with camps scattered up and down the railway for 30 miles, the lions could strike in a different camp each night.
It was not many days after Colonel Patterson arrived that reports started coming in about workers disappearing. Although it was told him that lions were responsible, Patterson at first didn't believe it. When he finally investigated, it was quickly (and gruesomely!) discovered that not one but two lions were responsible for killing the workers. Early attempts to shoot the lions were unsuccessful. The lions seemed to be able to predict what Patterson would do next. And, with camps scattered up and down the railway for 30 miles, the lions could strike in a different camp each nightThe workers soon constructed thorn bomas (A boma is a thorn fence erected around a dwelling or corral designed to keep predators such as lions out.) around their camps, and kept fires burning at night to try to scare off the lions. But it didn't work. The lions literally ignored the thorns, and would drag themselves and their meals right through them. This caused no small concern among the workers.Still, there were a couple of lucky escapes. One night, a lion attacked a man riding a donkey. The donkey was knocked over and the man knocked off. The lion moved in for the kill, and somehow got his claws hooked on a rope tied to some oil cans that had been around the donkey's neck. The lion couldn't immediately figure out how to unhook the rope, and the oil cans were making a terrible racket. The noise frightened the lion so much that it ran off back into the bush, dragging the oil cans with it. The rider escaped to the safety of a tree and stayed there the rest of the night.
Another time, one of the lions broke into a tent, and was intent on carrying off the occupant who was sleeping on a mattress. Instead, somehow, the lion got hold of the mattress and pulled it out from under the man. Soon realizing it's mistake, the lion dropped the mattress and ran off. Another time, one of the lions jumped onto a tent containing 14 Indian 'Coolies'. The lion broke through the tent, clawing up one man's shoulder in the process. Somehow in the ensuing confusion, the lion grabbed a sack of rice and made off with that instead. The lion 'threw it down in disgust' a short distance away and beat a hasty retreat.
At first, the effect of the lions grisly habits on the workers wasn't too great, because the camps were spread out over a large area. But, a large area. But, as the railhead pushed Northwestward, only a few hundred workers were left at Tsavo to build the bridge. The lions now directed their efforts on this one camp, and this is when the morale at Tsavo really began to drop! One night, one of the lions attacked the hospital tent. At first, he was scared away when the doctor's assistant knocked over a cabinet of supplies in fright. But, the lion tried again. Breaking through the tent, he seized one of the patients, and injured 2 of the other patients. It was decided after that to move the hospital tent. The very next night, a lion attacked the new hospital tent! Many of it's occupants got to witness the lion seize, kill and drag through the thorn boma the hospital's water-carrier. The next day, (as was usually the case for these lions) all that was left of the water-carrier was his head, a few of the larger bones, and part of his hand.The hospital-tent was moved again, and an even thicker boma was erected around it. A supply wagon was parked at the previous hospital tent location, and some cattle were tied up inside the boma as bait. Colonel Patterson and the Doctor stayed up all night, hoping to get a shot at the lions. Their vigilance was rewarded, as one lions jumped into the boma. Patterson and the doctor figured that the lion had silently dispatched one of the cattle and was trying to find a place to get out of the boma. Unbeknownst to them, the front entrance to the boma was not properly secured, and the lion exited by this route. It then proceeded to stalk Patterson and the Doctor. Luckily for them, they figured this out in time. The lion attacked, and Patterson got off a shot. This scared off the lion. Colonel Patterson writes much later in the Field Museum account of that incident that he believes that he shot one of the canine teeth out of the mouth of that lion. If this was indeed the case, this lion was the one now known as maneater #1. The date was April 23rd.
After the incident at the old hospital site, the lions left the area for a few months, and later accounts report people were disappearing at other camps some miles away. During this time, a considerable amount of effort was spent building an elaborate trap using a boxcar, to be ready should the lions return. The boxcar was divided into two compartments by strong bars. A trip wire was arranged to drop a gate made of railroad rails over the entrance to one of the compartments. (For lack of a suitable drill, Colonel Patterson 'drilled' holes in the rail sections with his .303 rifle!) There was a small, secure entrance into the other compartment. Thorns were piled all around the boxcar. For the first few nights, Colonel Patterson sat inside the secure compartment, acting as human bait. Should a lion sense his presence inside the boxcar, it would enter and trip on the trip wire. The gate would fall and trap the lion in the first compartment. The Colonel could dispatch the lion at his leisure from behind the strong bars. After a few days, he got some other people to take turns manning this trap.
The lions had been gone for so long that the workers started to let down their guard a little. This didn't last long. One night, some of the Coolies had decided to sleep outside their tent, but inside the boma. This would not be their lucky night. One of the lions jumped into the boma. Although stones and firebrands were hurled at the lion, it selected a victim, and dragged it out of the boma through the thorns! Outside the boma, the other lion joined the first one, and they enjoyed their meal not 30 yards away. Even though shots were fired at the lions, none of them made their mark, and the lions went right on eating.Soon, a sort of routine would settle in. The lions would be heard roaring in the vicinity of one of the camps (They were again spread out up and down the rail line.). Suddenly, they would stop roaring. Word would go from camp to camp, 'Beware brothers, the devil is coming!'. There would be agonizing shrieks from somewhere in the camp, and one less person at roll call the next morning.Every night, Colonel Patterson and others would stay up, hoping to get a shot at one of the lions. They never did.
The lions were getting bolder and bolder. Some nights, they would each take a victim, so they wouldn't have to share. They could go undetected right through the thorn fences. One night, a bunch of Coolies escaped the lions to the safety of a tree. They so heavily loaded the tree that it collapsed, throwing them to the ground very close to the lions. But, the lions didn't care. They had already caught a victim and were too busy feasting on him.
One night, the lions caught a victim and carried it close to Colonel Patterson's camp to devour. He vividly remembers the sound of bones being crunched and contented 'purring'. It took days to get these sounds out of his head. (Lions actually can't purr like smaller felines. They have a rough 'pseudo purr' they don't often use.) Finally, on December 1st, the workers approached Colonel Patterson and told him they would no longer be 'food for lions or devils'. As soon as this ultimatum was delivered, the workers went out and threw themselves down on the tracks in front of the next train. They then climbed on every available seat, and left Tsavo. The entire railway project ground to a halt. Only a small number of workers were brave enough to stay behind. These workers took up residence in trees, atop water in trees, atop water tanks, or in underground pits covered with logs.
On December 3rd, the Superintendent of Police arrived with twenty men to help hunt down the lions. It was on that night that one of the lions finally entered the rail-car trap. There were three armed men in the secure part of the trap. After the gate had fallen, the lion began to thrash at the bars, scaring the men silly. Finally, they got their courage up and began firing. Even though they could have touched the body of the lion with the muzzles of their rifles, nobody succeeded in getting a shot into it. Instead, one of the shots broke the chain holding the gate shut. The lion escaped with very minor injuries. The superintendent of Police, his men, Patterson and others tried to track the lions. Although one person got a brief glimpse, they again escaped! They tried again for two more days to track the lions. No success. The Superintendent and his men could not afford to stay any longer. Patterson was again left alone with the lions.
Patterson then rounded up his remaining Coolies, and instructed them to bring along things to make noise with. They surrounded the thicket. Patterson then stood at an animal trail coming out of the thicket. On his signal, the Coolies advanced, making all sorts of racket. It was then that Colonel Patterson actually saw one of the maneaters for the first time- a huge maneless male. He lifted his weapon to fire. Click! The twin-barreled rifle misfired! Intimidated by the noise, the lion jumped past Patterson and started to run off. He finally remembered he had another shot in his rifle and fired. He managed to hit the lion in the back, but it got away.
Dismayed, Colonel Patterson went back and looked at the donkey. The lion had just began it's meal on the hindquarters. (Lions like to start eating on the hindquarters because there is a lot of meat there without a lot of bones.) A nice donkey like this would be too good of a meal to abandon. The lion might return to finish his meal. Then, Patterson had an inspiration. Taking 4 poles, and a plank of wood, he constructed a platform in close to the donkey carcass. This platform, called a machan, would serve as an artifical 'tree' from which to hunt the lion. (There were no nearby trees to sit in.) He lashed the donkey carcass to a nearby tree stump with wire, so it couldn't be dragged off. Normally, Patterson would stand vigil with his gun-bearer (Colonel Patterson was an experienced big game hunter.), but tonight, he was sick. So, he began that night's vigil alone. It wasn't long before Patterson heard a twig snap and a sigh of hunger. The lion had indeed returned! But, as he watched, he determined the lion had detected him. Now, the lion was stalking Colonel Patterson! For two hours, the lion circled the rickety platform. All the lion would have to do is knock out a pole, or jump at the platform to dislodge Patterson. Instant easy meal! The whole situation made Patterson's flesh crawl.
Suddenly, something hit Patterson on the head! It turned out to be an owl that had confused him for a tree branch. He quickly recovered his wits. Below, the lion growled, and moved in for the kill! Very carefully, Patterson raised his rifle and fired. There was an angry growl, and the lion jumped around all over. He jumped into the bushes. Patterson fired away into bushes. The growls continued, but were growing weaker. They finally ceased altogether. One of the maneaters were finally dead!
Word quickly spread to the camps, and a wild celebration ensued. The next morning, the body of the lion was recovered. He was a maneless . He was a maneless male, 9 feet, 8 inches long from tip of nose to tip of tail. The lion had taken two shots-- one in the shoulder penetrating the heart (Probably the first shot), and another in one of the hind legs. It took 8 men to carry it to camp, and soon it was in the skinning shed The next morning, Colonel Patterson, his gun-bearer and a native tracker set off after the lion. The bloody trail was easy to follow. About a quarter mile away, they found him. He was hiding in the grass, glaring at the men with bared teeth.
Patterson aimed carefully, and took a shot. The lion then charged him with all the muster it had left. Another shot knocked him over, but he got up and continued his charge. A third shot had no effect. He reached for another rifle, only to discover his gun-bearer had abandoned him to the safety of a tree. Patterson had no choice but to follow suit. If one of the shots hadn't broken a hind leg, Patterson would have never made it. Once in the tree, the Colonel grabbed a carbine from his gun-bearer and shot the lion once more. The lion collapsed.
Rather foolishly, Patterson quickly climbed down. To his complete surprise, the lion charged again! A shot in the chest and another in the head (Look at the skull on the pictures page) finally did the brute in. Even so, the lion bit savagely at a branch until the last bit of life drained out of him.
Colonel Patterson had all he could do to prevent the dead lion from being torn apart by the workers. He had it carried to his nearby boma for examination. This lion (also a maneless male) had taken at least six shots. There was also a slug buried not far into the flesh of the back. This was the slug from the shot fired 10 days before. In any case, this lion measured 9 feet, 6 inches from tip of nose to tip of tail. Soon, this lion took it's turn in the skinning shed.
Thylacosmilus
First described in 1934 by Elmer Riggs, Thylacosmilus atrox was a large saber-toothed predator that lived and hunted on the plains of prehistoric South America. What is known about the creatures is derived mostly from two partial skeletons found in Pliocene deposits of Argentina, which up to now remain the most complete evidence ever found. The Thylacosmilus ('pouch-knife') and its prey lived and thrived during the late Miocene and early Pliocene and are believed to have succumbed to extinction around 2 million years ago. Often lumped into the category of other saber-toothed predators, mainly the "saber-toothed cats" of the true cat family, the Thylacosmilus atrox was neither a cat nor related to cats at all but rather a highly specialized species of prehistoric pouched marsupial. It was more closely related to the opossum and kangaroo than to any feline or other placental mammalian group.
TheThylacosmilus is an excellent example of parallel evolution. It had long dagger-shaped teeth that could stab through the thickest of hides. Out of all the saber-toothed puncture killers, the saber teeth of Thylacosmilus were the longest and most specialized of them all. The fangs were tapered to an edge on both the front and the back cusps. In addition, unique among the saber-toothed predators, the sabers continued to grow continuously throughout life to compensate for wear on the tips. These highly specialized features may have arisen as a countermeasure to the thick and sometimes armored coats that evolved among the contemporary herbivores that were its prey.
About the size and build of a modern jaguar, Thylacosmilus atrox was squat and powerfully built. Short, heavily muscled limbs indicates that it may have been an ambush hunter. Unlike its feline counterparts though, Thylacosmilus lacked retractable claws, an indication that it may have hunted in a very different way. Examination of the forelimbs shows an articulation and musculature that would have made it more than capable of grasping and subduing prey in a catlike manner.
Another similarity to the placental saber-tooths is the independent development of a long powerful neck. A long neck provides vertebrae with enlarged processes for muscle attachment. This provides for strong head-depressing musculature necessary for the stabbing action needed to successfully penetrate the hide and flesh of their prey.
Although anatomically similar in build and possibly in life-style as well, the Thylacosmilus would have looked uniquely distinct from other saber-toothed predators. They would have possessed marsupial characteristics not present in other animal groups. Color patterns, identifying marks, and social habits are indeterminable from fossil remains but may have parallels among modern marsupial predators. Such distinctions include subtle anatomical differences in the skulls of the marsupial saber-tooths compared to the skulls of their feline counterparts. The most distinct feature are the elongated flanges that jut down and gently slope inward from the lower jaw. These flanges would have acted like a natural sheath by serving to protect the teeth when the jaws were closed. The skull of Thylacosmilus atrox also possessed a postorbital bar, a very rare structure in carnivores that may have served as an area of attachment for temporalis fibers, and the aponeurosis, which overlays the entire muscle. It would have also shielded and secured the eyes from pressure by the contracting jaw muscles.
Thylacosmilus atrox would have reproduced in the same way as all marsupials, nursing their young to maturity by way of an external pouch as seen in kangaroos and wombats. The pouches may have opened to the back as seen in the recently extinct thylacines. Such an adaptation would have served to better protect the young while the mother raced through brush in pursuit of prey.
Thylacosmilus and related species are believed to have evolved into hunters from primitive marsupial insectivores that became isolated when South America drifted away from the rest of the continents. Isolation allowed marsupial as well as placental evolution to follow unique paths and fill unexploited niches left open after the demise of the dinosaurs. The result was many diverse, geographically distinct creatures that existed nowhere else on Earth. Thylacosmilus and their type are collectively known as borhyaenids ['meat/food-hyaenids']: the lions, tigers, and bears of their time. The doglike borhyaenids and saber-toothed thylacosmilids are placed into separate families, the Borhyaenidae and Thylacosmilidae respectively, within the superfamily Borhyaenoidea.
Thylacosmilus and its relatives survived unchallenged until around 2 million years ago when they ultimately became extinct. There are a couple theories attributed to their demise. The first of the two describes a great upheaval that completely changed the makeup of South American fauna. The upheaval followed a temporary end to the continent's isolation, when sea levels dropped and a land bridge emerged to join North and South America. Highly adapted mammals from the north would have migrated southward across the newly formed bridge. This "northern invasion" may have introduce new types of predators such as jaguars and pumas, which may have pressured and competed with the Thylacosmilus to the point of extinction. Another theory also suggests that the Thylacosmilus was outcompeted, not by other mammals though but by large, fast-running ground birds that used massive beaks to stab and rip their prey. These ground birds known as phororhacoids ('thief-rags-like', also phorusrhacoids) were more like latter-day carnivorous dinosaurs than birds. Their power and speed may have proved superior to the thylacosmilids and borhyaenids, winning out in the role for top predators. More recently there has come to light a new theory that suggests that the Thylacosmilus and other contemporary fauna may have fallen to extinction as a result of a meteor impact, much in the same way as the dinosaurs. Although it would have been on a much smaller scale, a localized impact could have upset the ecosystem causing the demise of entire groups of animals, especially if the species were geographically distinct to the blast area. Evidence for this is based on a stratum of rock and sediment containing natural glass formed of sand from intense heat. The intense heat is believed to have occurred as a result of a meteor impact, with the logic being that below the glass layer fossils of Thylacosmilus and other contemporary animals can be found but not above the layer, an intriguing theory. Clearly more evidence and further study is needed, and an exact cause for the extinction may never be known.
TheThylacosmilus is an excellent example of parallel evolution. It had long dagger-shaped teeth that could stab through the thickest of hides. Out of all the saber-toothed puncture killers, the saber teeth of Thylacosmilus were the longest and most specialized of them all. The fangs were tapered to an edge on both the front and the back cusps. In addition, unique among the saber-toothed predators, the sabers continued to grow continuously throughout life to compensate for wear on the tips. These highly specialized features may have arisen as a countermeasure to the thick and sometimes armored coats that evolved among the contemporary herbivores that were its prey.
About the size and build of a modern jaguar, Thylacosmilus atrox was squat and powerfully built. Short, heavily muscled limbs indicates that it may have been an ambush hunter. Unlike its feline counterparts though, Thylacosmilus lacked retractable claws, an indication that it may have hunted in a very different way. Examination of the forelimbs shows an articulation and musculature that would have made it more than capable of grasping and subduing prey in a catlike manner.
Another similarity to the placental saber-tooths is the independent development of a long powerful neck. A long neck provides vertebrae with enlarged processes for muscle attachment. This provides for strong head-depressing musculature necessary for the stabbing action needed to successfully penetrate the hide and flesh of their prey.
Although anatomically similar in build and possibly in life-style as well, the Thylacosmilus would have looked uniquely distinct from other saber-toothed predators. They would have possessed marsupial characteristics not present in other animal groups. Color patterns, identifying marks, and social habits are indeterminable from fossil remains but may have parallels among modern marsupial predators. Such distinctions include subtle anatomical differences in the skulls of the marsupial saber-tooths compared to the skulls of their feline counterparts. The most distinct feature are the elongated flanges that jut down and gently slope inward from the lower jaw. These flanges would have acted like a natural sheath by serving to protect the teeth when the jaws were closed. The skull of Thylacosmilus atrox also possessed a postorbital bar, a very rare structure in carnivores that may have served as an area of attachment for temporalis fibers, and the aponeurosis, which overlays the entire muscle. It would have also shielded and secured the eyes from pressure by the contracting jaw muscles.
Thylacosmilus atrox would have reproduced in the same way as all marsupials, nursing their young to maturity by way of an external pouch as seen in kangaroos and wombats. The pouches may have opened to the back as seen in the recently extinct thylacines. Such an adaptation would have served to better protect the young while the mother raced through brush in pursuit of prey.
Thylacosmilus and related species are believed to have evolved into hunters from primitive marsupial insectivores that became isolated when South America drifted away from the rest of the continents. Isolation allowed marsupial as well as placental evolution to follow unique paths and fill unexploited niches left open after the demise of the dinosaurs. The result was many diverse, geographically distinct creatures that existed nowhere else on Earth. Thylacosmilus and their type are collectively known as borhyaenids ['meat/food-hyaenids']: the lions, tigers, and bears of their time. The doglike borhyaenids and saber-toothed thylacosmilids are placed into separate families, the Borhyaenidae and Thylacosmilidae respectively, within the superfamily Borhyaenoidea.
Thylacosmilus and its relatives survived unchallenged until around 2 million years ago when they ultimately became extinct. There are a couple theories attributed to their demise. The first of the two describes a great upheaval that completely changed the makeup of South American fauna. The upheaval followed a temporary end to the continent's isolation, when sea levels dropped and a land bridge emerged to join North and South America. Highly adapted mammals from the north would have migrated southward across the newly formed bridge. This "northern invasion" may have introduce new types of predators such as jaguars and pumas, which may have pressured and competed with the Thylacosmilus to the point of extinction. Another theory also suggests that the Thylacosmilus was outcompeted, not by other mammals though but by large, fast-running ground birds that used massive beaks to stab and rip their prey. These ground birds known as phororhacoids ('thief-rags-like', also phorusrhacoids) were more like latter-day carnivorous dinosaurs than birds. Their power and speed may have proved superior to the thylacosmilids and borhyaenids, winning out in the role for top predators. More recently there has come to light a new theory that suggests that the Thylacosmilus and other contemporary fauna may have fallen to extinction as a result of a meteor impact, much in the same way as the dinosaurs. Although it would have been on a much smaller scale, a localized impact could have upset the ecosystem causing the demise of entire groups of animals, especially if the species were geographically distinct to the blast area. Evidence for this is based on a stratum of rock and sediment containing natural glass formed of sand from intense heat. The intense heat is believed to have occurred as a result of a meteor impact, with the logic being that below the glass layer fossils of Thylacosmilus and other contemporary animals can be found but not above the layer, an intriguing theory. Clearly more evidence and further study is needed, and an exact cause for the extinction may never be known.
Prehistoric
Although we are familiar with cinema representations of sabre-toothed cats, only a handful of prehistoric big cats exceeded an average weights of 100 kilos and only four or five North American prehistoric big cats (not all are true cats) are in the 100+ kilos category. This means few were longer or taller at the shoulder than a modern leopard or jaguar. Many of the "bigger" prehistoric big cats were compact, muscular animals. Modern big cats are relatively long-legged and lithe in comparison.
Although they are often called "big cats" many of the prehistoric species were not true cats, but were cat-like mammals. South America and Australia were both island continents and lacked placental mammals. The "lions" and "tigers" of those continents were lion-like marsupials, more closely related to kangaroos, wombats and their like. Theyare known as "cat analogues" as they resembled cats and filled the same ecological niche as cats. Another line of prehistoric cats were the Paleofelids ("ancient cats") that developed in parallel with the true cats and from a common ancestor, but which have left no modern descendants. Finally there are the prehistoric true cats, extinct relatives of modern cats. Why would evolution create creatures very similar in form to cats? It's an example of convergent evolution - there are a limited number of solutions to environmental conditions hence animals that aren't closely related often independently evolve similar traits as they both adapt to similar environments and niches. The cat-like form has evolved at least three times: marsupial lions/tigers, Paleofelids and Neofelids. Sabre-toothed cat-like animals evolved separately four times in evolutionary history: Nimravids, Felids, Creodonts and the Thylacosmilids.
The taxonomy (classification) of both living and extinct species changes frequently. As more fossil evidence comes to light, species are reclassified. Some are given their own species or genus while others are absorbed into an existing species or genus and their original classification is scrapped.
Although there are references to animals being found in places far apart on the modern world map, the continents used to look very different. Some land masses that were once joined together have now split and drifted apart, others that were far apart have collided. Some land masses that are currently not joined to each other were joined by ancient land bridges when sea levels were much lower than they are today.
Carnivorous mammals evolved from Miacids small pine marten-like insectivores that lived 60 million - 55 million years ago. The miacids split into two lines: Miacidae and Viverravidae. Miacidae gave rise to Arctoidea/Canoidea group (bears and dogs) while Viverravidae gave rise to Aeluroidea/Feloidea group (cats, hyenas, civets, mongooses) around 48 million years ago. The Viverravidae also gave rise to a group called Nimravidae. The Nimravids were cat-like creatures that evolved in parallel with true cats; they are not part of true cat lineage and have left no living descendents.The first true cat to arise from Viverravidae was Proailurus (first cat") around 30 million years ago. The best-known species was P lemanensis, found in France.Proailurus was a small weasel-like cat with relatively short legs and a long body. It had one more premolar on each side of its bottom jaw than do modern cats. About 20 million years ago, Proailurus gave rise to Pseudaelurus. Pseudaelurus were Miocene ancestors of cats. Pseudaelurus lorteti was about the size of a large lynx while P validus was the size of a large lynx or small puma. Three other species of early cat are described as Pratifelis, Vishnufelis and Sivaelurus (S chinjienis).Pseudaelurus went on to split into two major groups: the Machairodontinae (true sabre-tooths) and Schizailurus (the ancestor of the modern day Felidae group).
18 million years ago, Schizailurus gave rise to the Felidae. The first of the modern Felids were the early cheetahs; now represented by Acinonyx (modern cheetah); true cheetahs are believed to have evolved around 7 million years ago. Some sources claim Miracinonyx (North American cheetahs) evolved only 4 million years ago from Acinonyx, but recent studies show Miracinonyx was probably ancestral to both cheetahs and puma and was intermediate in type between these two modern species.
Around 12 million years ago, genus Felis appeared and eventually gave rise to many of our small cats. Two of the first modern Felis species were Felis lunensis (Martelli's cat, extinct), and Felis manul (Manul or Pallas's Cat, living). Extinct Felis species are: F attica, F bituminosa, F daggetti, F issiodorensis (Issoire Lynx), F lunensis and F vorohuensis. The ancestor of modern Felis species was F attica. Genus Panthera ("biting cats" or "roaring cats") genera evolved around 3 million years ago; there are a number of extinct species discussed later in this article.
Genera Acinonyx, Felis and Panthera are all represented today and taxa of some modern species is regularly revised as more complete fossils of ancestral species are found, giving a clearer indication of who begat whom and when various lineages split.
The jaguar-sized Thylacosmilus ("pouched blade") was a large, predatory marsupial; part of a unique group of predators on the South American pampas; the borhyaenids. These appeared in the Miocene and ruled the South American forests and woodlands for some 30 million years, but have no known ancestor or descendant. Thylacosmilus was the most successful member of that family and was the ultimate mammalian predator of its day in Plio-Pleistocene South America, but when the continents of North America and South America joined, it lost out to the more highly developed and faster eutherian cats.
Two species are described: T atrox and T lentis. Thylacosmilus atrox looked like a sabre-toothed cat, but is more closely related to kangaroos. As far as we know, Thylacosmilus was the only marsupial to have developed the sabre-toothed weapon. Like Smilodon, the eutherian sabre-tooth tiger, it had adapted to hunting mega-fauna.
Thylacosmilus lacked incisor teeth, but had very long upper canine teeth that grew continually. These long stabbing teeth projected below the mouth-line. Strong neck and jaw muscles allowed the sabre-teeth to be driven downward with a tremendous killing force. Its huge stabbing teeth were about 15 cm (6 inches) long (longer than those of Smilodon) and may have been used to slash the soft throat of its prey. The jaws were capable of a gape that left the teeth clear to do their work. These sabres grew continually throughout Thylacosmilus's life, much like the incisors of modern rodents. Unlike Smilodon (see later), it had no scabbard-like tooth-guards on its lower jaw though its skull had a deep flange on its lower jaw, forming a protective sheath for when the sabre teeth were not being used.
Unlike modern cats, which tend to be sleek and long-legged, it appears to have been short-legged and heavily built, being about 1.2 metres (4 ft) long and weighing around 100 kilos. Its claws were not retractile. It probably preyed on large, slow-moving mammals and when the two continents joined, the highly specialised Thylacosmilus could not compete against the faster, sleeker eutherian big cats. South America has also had at least three species of cats whose body weights exceeded 300 kilos - about twice the weight of modern lions.
The Thylacoleonidae were lion-like marsupials that inhabited Australia in Oligocene to Pleistocene times. They probably hunted across the Australian grasslands, although some may have been arboreal. They were vombatomorphian (wombat-like) marsupials, evolved from herbivore ancestors; their closest living relatives being koalas and wombats. The more primitive species had generalised crushing molar teeth (like modern omnivores) as well as carnassial blades. In more specialised species, the crushing molars were reduced or absent and the carnassials had become huge.
The Thylacoleonidae ranged from the size of a domestic cat to the size of a leopard and possibly even the size of a lion (1.7 metres/5 ft 6 in). So far, eight species of marsupial lion have been discovered and there may be at least two more. Those of genus Wakaleo were leopard-sized and designed for power rather than speed. W alcootaensis was slightly larger than W oldfieldi or W vanderleueri. These "marsupial leopards" may have ambushed prey from tree branches. Priscaleo was much smaller. P pitikantensis was about the size of a modern Australian possum. P roskellyae was about the size of a domestic cat, possibly up to ocelot-sized, and may have been arboreal.
The most famous member of this family is Thylacoleo carnifex, the "marsupial lion". This was Australia's equivalent to the South American marsupial Thylacosmilus atrox and to the eutherian Smilodon. Its enormous meat-shearing carnassial (cheek) teeth were the largest of any mammalian predator. It also had bolt-cutter incisors, and switch-blade-like claws on its semi-opposable thumbs. It was the most specialised mammalian carnivore ever known; entirely lacking grinding teeth. Because T carnifex lacked large canines, it was originally believed to be a herbivore, using its unusual front teeth and claws to break open nuts and fruit; its lack of grinding teeth suggest a diet of soft fruit such as melons! However, wear on the teeth indicates a meat-eating diet, and it probably preyed on giant kangaroos and wombats of the time. Compared to sabre-tigers, such as Smilodon, it had a short cat-like face and more elaborate carnassial teeth, giving it a powerful killing bite. Most modern cats have carnassials that can crunch smaller bones, but Thylacoleo's teeth lacked bone-crunching adaptations and were entirely adapted to shearing soft tissue. Projecting front incisors were modified into killing teeth, and looked rather like the canines in the placental carnivores; the actual canine teeth were insignificant.
T carnifex had a short body, closer in length to that of a leopard rather than a lion, but the bones of its legs show it was far more robust than a leopard. Estimates derived from size (partly based on skull size) and robustness suggest it weighed between 100 and 130 kilos, putting it in the same size range as modern tigers and lions. It was extremely robust and built for power rather than endurance, with tremendously powerful forelimbs. It probably ambushed prey as large as, or larger than, itself, using the thumb claws to hold the prey in a deadly embrace while employing its fang-like incisors.
T carnifex survived until around 50,000 years ago and may have come into conflict with early Aboriginal settlers entering Australia. Conflict and competition with humans, and with the introduced dingo, may have contributed to the extinction of this highly specialised carnivore. There are theories that relict populations of smaller marsupial lions survive in the form of the cryptozoological "Queensland Tiger". Like Thylacoleo, the Queensland Tiger is described as short-headed, sharp-clawed and superficially cat like. Eyewitnesses (and a single photograph) show it to have vertical striped on the forequarters. It has never been positively identified.
The Nimravidae were a separate family of cat-like animals that evolved parallel to the true cats (Felidae). The common ancestor of the Nimravidae and the Felidae was the Viverravidae (feline-like) group of miacids some 55 million years ago in the late Eocene. Proailurus, a descendent of the miacids, evolved into Pseudaelurus, which split into two main groups, one of which became the modern cats (Neofelids), and one became the Machairodonts (true sabre-toothed cats, a sub-family of the Felidae). Nimravids are also known as the Paleofelids (ancient cats), or False Sabre-toothed Cats (because they are cat-like, but not true cats). They were the earliest cats to evolve and lived from the late Eocene (36 million years ago) to the late Miocene (5 million years ago), peaking around 28 million years ago. The three subfamilies of nimravid and their genera are shown in the table below. Some are known from single or fragmented specimens and the taxonomy may change as more fossils are discovered.
Nimravids had different skulls to true cats. The structure of their middle and inner ear was different and many Nimravids had a flange on the front of the lower jaw (also seen in some machairodonts [true sabre-tooths] and thylacosmilids [marsupial sabre-tooth]). The flange is a bony prominence that projects downwards and is as long as the canine teeth; the teeth fit into a groove. Barbourofelis has the most prominent flange, while Nimravus and Dinaelurus lacked a flange. The actual sabres were narrow, pointed canines whose length varied according to species.
Nimravids were very cat-like in appearance and had retractile claws. Many were muscular and low slung with heavy-set bodies on short legs. Like the modern lynx, some were short-tailed although many others were long-bodied and long-tailed. The skulls of genera Dinictis, Nimravus and Dinaelurus are especially cat-like. Their prominent upper canines were longer than those of modern cats, but shorter than those of the true sabre-toothed cats; their lower canines were proportionally longer. The most common species in the fossil record are those belonging to Dinictis, Eusmilus, Hoplophoneus and Barbourofelis.
Hoplophoneus lived during the late Oligocene (33 - 30.5 million years ago), some 20 million years before Smilodon. Some earlier authors erroneously place Hoplophoneus among the Felidae (true cats) as the ancestor of Smilodon and true sabre-tooths, but current fossil evidence makes this incorrect and indicates that Hoplophoneus and Smilodon are from different evolutionary lines. Some were the size of bobcats while others were jaguar-sized. Like many other Nimravids, Hoplophoneous had a bony flange into which its curved canines fit.
Nimravus has been found in France and parts of North America from the early Oligocene to early Miocene. Some were 1.2 metres (4 ft) long. With its sleek body, it may have resembled the modern caracal, although it had a longer back and more dog-like feet with partially retractile claws. It competed with other false sabre-tooths such as Eusmilus. A Nimravus skull, found in North America, had been pierced in the forehead region, the hole exactly matching the dimensions of Eusrnilus' sabre tooth; Nimravus survived as the wound showed signs of healing. It probably hunted birds and small mammals, ambushing them like modern cats, rather than chasing them down. Some specimens still have unclear classification, for example a species once identified as N catacopis is closer to true cats and is now classified as Machairodus aphanistus (previously Machairodus catacopis).
Dinictis was a small nimravid that lived on the plains of North America during the late Eocene and early Oligocene (40 million years ago). Dinictids had a sleek bodies, short legs, long tails, and walked plantigrade ("on the whole foot" modern cats walk digitigrade "on the toes").
Eusmilus was a dirk-toothed cat found in France and parts of North America during the late Oligocene (30.5-28.5 million years ago). It was noted for its long, flattened sabres and very prominent mandibular flange. Most were leopard-sized and rather long-bodied and short-legged compared to modern leopards. Some reached 2.5 metres (8 ft) long. It was a typical false sabre-tooth with enlarged upper canines, but insignificant lower canines, while many of the other teeth had been lost to accommodate its sabres (Eusmilus had 26 teeth, compared to 44 teeth in other carnivores). The jaw hinge was modified to open to an angle of 90 degrees to allow the great sabre teeth to do their work. Its lower jaw had bony guards that lay along the length of the sabres, protecting them from damage when the mouth was closed. There is fossil evidence of conflict between Eusmilus and Nimravus.
The lion-sized Barbourofelis was one of the most recent members of the Nimravidae family. It lived during the late Miocene (15 million - 6 million years ago) and had the longest canines of all the nimravids. They had very prominent flanges on the lower jaws and an unusually shaped skull. The Barbourofelids were probably very muscular, resembling a bear-like lion or lion-like bear. Pogonodon was a cat-like sabre-tooth.
The Machairodontinae are true cats and their fossils have been found in North America, Europe, Asia and Africa. Although we tend to think of the sabre-toothed tiger, there were two varieties of sabre-toothed cats: dirk-toothed cats and scimitar-toothed cats. Dirk-toothed cats had two long, narrow upper canines, and were usually short-legged and stocky. Scimitar-toothed cats had upper canines that were shorter and broader, longer, thinner legs and were generally more lithe. The exception was a cat known as Xenosmilus, which has the short, broad canines of a scimitar-toothed cat, but has short legs.
Modern cats have conical canine teeth, but the machairodonts' (machairodont means "sabre tooth") canines were flattened from side to side (like a blade) as well as being elongated. To accommodate their large canines, they had fewer upper premolar teeth. Their incisor teeth were larger, angled differently and placed further forward than in modern cats. Other adaptations allowed them to open their jaws extremely wide and gave them strong neck muscles. Some species, such as Megantereon, had a bony flange that protruded downward from the front of their lower jaw.
Although the sabre-toothed cats have long dagger-like canines, these were probably too blunt and fragile to be used to stab prey. They were unlikely to have gone for the nape of the neck to sever the spine, like many modern cats. If they hit bone, they could shatter (leading to abscesses and possibly fatal bacterial infections). The current theory is that sabre-toothed cats went for the soft throat of their prey, using their powerful teeth to sever the arteries and windpipe.
These were Eurasian scimitar-toothed cats that lived during the late Miocene to early Pleistocene.
The best known genus is Dinofelis ("giant cat") which lived in Eurasia, Africa and North America around 5 million - 1.5 million years ago. In South Africa, Dinofelis has also been found at sites along with Australopithecines, so it was probably a predator of our own ancestors. Genus Dinofelis includes cats previously classified as Therailurus. It became extinct in Eurasia and North America during the Early Pleistocene, but survived in Africa until the Mid-Pleistocene. The largest known form was the Chinese D abeli. Its size and build are comparable to a large leopard or jaguar (1.2 metres/4 ft) with forelimbs more heavily built than hind-limbs. Like the modern jaguar, they may have been forest-dwellers. Dinofelis ("giant cat") had flattened canines considerably shorter than those of the sabre-tooths, scimitar-tooths or even dirk-tooth cats. The canines were longer than those of biting cats (those that kill prey with a single neck-bite) so it is debatable as to which subfamily of the Dinofelis belongs.
Metailurus lived 15 - 8 million years ago and were similar to Dinofelis in body type and shape. They were smaller than Dinofelis, being leopard sized, but had longer canine teeth, intermediate between modern cats and sabre-toothed cats. Like Dinofelis, Metailurus was stockily built with forelimbs more heavily built that its hind limbs. Its build suggests it hunted in forested areas and may have been more arboreal than Dinofelis.
The puma-sized Adelphailurus is known from a single type specimen from Kansas, USA. Unusually for a cat, Adelphailurus kansensis retained the upper second premolar, a trait it shared with Stenailurus. The members of genus Pontosmilus were formerly classified as Paramachairodus.
This comprises the genera Machairodus, Homotherium and Xenosmilus. Classification is liable to be revised as more fossils are discovered and there are museum specimens assigned to 4 other genera.
Machairodus is a variable genus of large sabre-toothed cats ranging in size and structure from smaller varieties right up to lion-sized. They were found in Europe, Asia, Africa and North America from 15 - 2 million years ago. The taxonomy is debatable. They have large canine teeth, and its incisors, canines and carnassials has serrated edges. There were two basic types of Machairodus: the primitive type and the evolved type (possibly adaptations to different environments). The more primitive types included M aphanistus and resembled Smilodon. The more evolved type had serrated teeth and elongated forelimbs structurally similar to the hyena-like Homotherium; they may have been ancestral to Homotherium. The variable forms indicate a adaptations to different environments ranging from forest/woodland dwelling to plains hunting. The hyena-like species may have covered long distances while hunting or for opportunistic scavenging.
Homotherium (therium = "beast") is a group of unique hyena-like sabre-toothed cats that also ranged widely (Africa, Asia, Europe, North America) from 3 million - 0.5 million years ago. They were about 1.2 metres (4 ft) long with front limbs longer than the rear ones. Homotherium's incisors were very large and robust and they had serrated medium-length canine teeth. Homotherium is a scimitar-toothed cat i.e. it has shorter, flatter canines than other sabre-tooth cats and its canines curve backwards like scimitar blades. Homotherium would have had the sloping look of a hyena with slender legs and relatively long neck. Its anatomy suggest that it walked with the whole foot on the ground (plantigrade) like a bear. This hyena-like conformation may have allowed them to cover long distances when hunting. It is more likely to have walked semi-plantigrade, the back sloping slightly; an adaptation for greater strength.
Hometherium survived until the end of the last ice age about 14,000 years ago and probably preyed on mammoths, possibly hunting in family groups. In Texas, the bones of a family group of scimitar-tooths are preserved alongside young mammoths and their eventual extinction was probably linked to a decline in prey species. As an adaptation to ice age conditions, some species may have been white or pale grey (like modern arctic predators).
H serum, the North American scimitar cat ,was originally named Dinobastis serus. It was short-tailed and slender-limbed, with relatively long forelimbs and short, powerful hindlimbs. Its deepened chin meant that its upper canines did not protrude beyond the lower margin of the lower jaw. H serum's large nasal opening, like that of the cheetah, would have allowed quicker oxygen intake aiding in rapid running. Skulls show it had a large visual cortex, indicative of a daytime hunter. It was built for short bursts of speed, rather than long chases. The claws of its forelimbs were not retractile, allowing better traction at high speed. Its hind limbs were shorter than its forelimbs and had a bear-like heel and ankle. The long hind feet had non-retractile claws.
H latidens is depicted in paleolithic stone carvings from Isturitz, south-western France show a short-tailed big cat with a deeply set lower jaw. This matches the traits of the European Scimitar-tooth, H latidens. The carving suggests that the cats had spotted pelts and paler undersides. H ultimum, the Asian scimitar cat, occurred in China.
Homotherium ischyros (or Ischyrosmilus), had canines serrated like steak-knives, along their front and back edges. This made it easier to slice through the skin of thick-skinned prey. Megantereon lacked these serrations on its upper canines. Ischyrosmilus's exact taxonomy is unclear and it may be one of the Smilodontini.
Xenosmilus ("strange knife") lived in the Florida, USA region 1.7 - 1 million years ago and was a robust cat with short legs and very thick, stout canine teeth. It was lion-sized, very robust and somewhat bear-like Pleistocene felid (length 2 metres / 7 ft, weight 180-230 kg / 400-500 lbs). Like Homotherium, it had broad, knifelike, coarsely crenulated teeth and projecting long, curved, serrated incisors, but it had the short, stout-legged features of Smilodon. Originally palaeontologists thought they had mixed up the bones of 2 other species since dirk-tooth cats were bear-like with two long, narrow upper canine teeth and short legs, while scimitar-tooth cats were longer-legged with two shorter, broader upper canines. Xenosmilus had a mix of features: the short, broad coarsely crenulated teeth of a chasing cat, but the stocky legs of an ambush hunter. It probably stalked close to its prey and then sprinted from cover to catch it. The specimens were found with bones of peccaries (wild pigs), giving an indication of its main prey. It appears to have been a more specialised sabre-toothed cat than Smilodon and its size made it the most ferocious sabre-tooth in the world at the time. Smilodon may only have become a dominant predator after Xenosmilus vanished.
This contains the famous sabre-toothed "tiger", Smilodon ("blade tooth"), of the late Pleistocene age (1.5 million - 10,000 years ago). The three known species were widespread throughout the Americas. Smilodon was stocky, with short, powerful legs and a bobtail. Their canine teeth were the longest of all the true sabre-toothed cats, being about 18 cm long. The South America Smilodon populator was the largest species. The South American Smilodon neogaeus is probably a synonym for S populator. The more famous Smilodon fatalis was found across North and South America, having migrated there from North America during the Pleistocene. S fatalis is sometimes divided into two separate species: S californicus and S floridanus (these may have been sub-species or variant populations i.e. S fatalis californicus and S fatalis floridanus). They are usually compared to the modern lion based on body length (1.2 metres/4 ft) and conformation, but were more robustly and powerfully built. S fatalis and S populator were around 11/2 - 2 times heavier than the average lion (around 170 kilos). Smilodon gracilis was the smallest of the Smilodons and the earliest species (about 2.5 million years ago); it was found in the eastern United States and weighed around 80 kilos.
Smilodon's anatomy shows them to be were specialised hunters of big game; they probably ambushed their prey - their robust build is not designed for chasing it down. Most modern cats have carnassials that can crunch smaller bones, but Smilodon's teeth lacked bone-crunching adaptations and were entirely adapted to shearing soft tissue. The muscles of its shoulders and neck were arranged to produce a powerful downward lunge of its massive head. The jaw opened to an angle of over 120 degrees, to allow the huge upper canines to be driven into prey. The canines were oval in cross-section to retain strength, but also to ensure minimum resistance as they were sunk into the prey. They were also serrated along their rear edges, so they pierced flesh more easily. They probably preyed on large, slow-moving, thick-skinned herbivores, but also scavenged dead and dying animals. More than 2000 Smilodon skeletons have been recovered from the Pleistocene tar pits of La Brea (Los Angeles, USA) where they had been fatally lured by large animals trapped in the tar.
It seems likely that Smilodon lived in family groups, much like modern lions, and possibly hunted in groups. Some specimens showed signs of healed fractures, suggesting they ate food caught by family members. Their extinction seems to have coincided with open plains taking over from forest/woodland. Smilodon was not built for the chase and this reduction in cover would have made it harder to ambush prey.
Megantereon was another genus of cats with impressive canine teeth, although they are not well-represented. About the size of a jaguar (1.2 metres/4 ft), they flourished in the Mediterranean and spread throughout Africa, Eurasia and North America around 3 million - 1 million years ago. The only complete skeleton was found in France. They had very large upper canines but relatively small lower ones. Though impressive, its teeth were more like daggers than sabres in size and shape, hence Megantereon and its immediate relatives are referred to as dirk-tooth (dagger-tooth) cats. They are also characterized by their prominent mandibular flange. Only a single species, Megantereon cultridens, is known from fossil records, although some Chinese specimens are known as M. nihowanensis. They are believed by most to be the direct ancestors of Smilodon and other later sabre-tooth cats.
Also in this group is Paramachairodus, although it is under much debate as to the placement of this genus. Many animals formerly placed in this genus have been reassigned to Pontosmilus and placed within Metailurini. They are thought to have existed between 20 - 9 million years ago. The two species known are Paramachairodus ogygia (Spain) and Paramachairodus orientalis, plus the disputed P maximiliani..
Modern Felids evolved around 18 million years ago. The first of these were the early cheetahs; now represented by Acinonyx (modern cheetah); true cheetahs are believed to have evolved around 7 million years ago. Some sources claim Miracinonyx (North American cheetahs) evolved only 4 million years ago from Acinonyx, but recent studies show ("miracle cheetah") was probably ancestral to both cheetahs and puma and was intermediate in type between these two modern species. Another cheetah was the Sivapanthera genus.
Cheetah-like cats arose around 18 million years ago. According to some studies, the ancestor of modern cheetahs originated in Africa during the Miocene and later migrated, giving rise to the now-extinct North American cheetahs. More recent studies suggest that a North American cheetah called Miracinonyx was the ancestor of both African cheetahs (modern Acinonyx) and American pumas (Puma concolor). Miracinonyx would have migrated across continents during the Ice Age. Miracinonyx inexpectatus [M studeri] existed in North America during the early Pleistocene (1 to 1.5 millions years ago) and may be even older. It had proportions intermediate between the modern cheetah and modern pumaAs a result, iIt is sometimes linked to the cryptozoological Onza (a gracile form of puma). Two species of cheetah inhabited late Pleistocene North America (100,000 years ago): Miracinonyx inexpectatus [M studeri] and Miracinonyx trumani.
Fossil evidence of early cheetahs is fragmentary, but Miracinonyx resembles modern cheetahs in having a short face, wide nasal passages and long body, but were less lanky. M inexpectatus and M trumani may be the reason North American evolved into such fast sprinters; North America has no living predator able to match the pronghorn's in speed. Unlike modern cheetahs, Miracinonyx inexpectatus had fully retractile claws and more robust conformation with shorter limbs than modern cheetahs. M inexpectatus would have been faster than the puma, but not as accomplished a sprinter as modern cheetahs; it was also better equipped for climbing.
The early true cheetah, Acinonyx pardinensis, appeared during the Pliocene and at 200 lbs were much larger than modern cheetahs. Known as Giant Cheetahs, they became widespread in China, southern Europe and India throughout the Ice Age, were lion-sized cheetah and probably as fast as modern cheetahs. Intermediate-sized cheetahs, Acinonyx intermedius, ranged from Africa as Far East as China during the mid-Pleistocene and became adapted to hunting on open grassland. These were larger than modern cheetahs. Acinonyx parchidinensis was the Pleistocene cheetah. The smaller modern cheetah, A jubatus, was once much more widespread, but became extinct in eastern Asia at the end of the Ice Age.
Biting cats are so-called because their relatively short, strong canines are adapted to dispatching prey by biting the bones and sinew of the neck and throttling it. Genus Panthera genera evolved around 3 million years ago and these have become the modern day big cats ("roaring cats" or "biting cats"). Prehistoric relatives of modern big cats lived between the Pleistocene to Recent times and ranged across South Africa, Asia, Europe and North America. Some, such as the "cave lion" were truly impressive creatures, reaching 3.5 metres (111/2 ft). The modern lion, Panthera leo, is now restricted to parts of Africa and to western India. There are extinct sub-species of the modern lion; until recent times there were sub-species in Arabia and Iran (the Barbary lion has been rediscovered and is being conserved).
Primitive lions (P leo fossilis) dispersed in the Old World about 500,000 years ago. P youngi, from north-eastern China 350,000 years ago, had similarities to both P leo spelaea (European Cave Lion) and P leo atrox (North American Cave Lion) and may be a link between the Eurasian/American cave lions and Asian/African modern lions. Extinct Panthera species include: P atrox (P leo atrox) (American lion), P gombaszoegensis (European jaguar), P leo spelaea (European Cave Lion), P palaeosinensis (a primitive leopard), P pardoides (a primitive leopard), P schaubi (a short-faced leopard) and P toscana (Tuscany lion, Tuscany jaguar). Some of these classifications are being revised as more complete specimens are discovered. Panthera schaubi (syn Viretailurus schaubi), which resembles a short-faced leopard, is currently regarded as an Old World puma, Puma schaubi.
Panthera leo spelaea, the European Cave Lion (300,000 to 10,000 years ago), was probably the largest cat that ever lived, being around 25% larger than modern lions, and larger than modern Siberian tigers. It was probably comparable in size to modern liger hybrids. It ranged across most of Europe and is depicted in cave paintings. Cave paintings and remains show that it lived until historical times in south-eastern Europe and possibly as recently as 2000 years ago in the Balkans. Cave paintings from Germany show cave lions as having ruffs or manes and tufted tails. A wall engraving from France and an ivory figurine of a lion-pelt-wearing human from Russia indicate faintly striped pelts. European Cave Lions inhabited steppes and parkland regions in the north and semi-desert areas in the south of Eurasia. They were evidently not adapted to deep snow or to dense forests.
Panthera atrox (P leo atrox) was a North American lion whose range extended to northern South America (Peru). P atrox crossed to North America over the Bering Strait land bridge during the last ice age, about 35,000 to 20,000 years ago. Its remains have been found in Alaska and some specimens have been found in the La Brea tar pits, Los Angeles, USA. Relatively few P atrox fossils have been recovered from La Brea compared to fossils of other carnivores; this lion may have been intelligent enough to avoid the natural traps. It probably hunted deer, North American horses (which became extinct and were reintroduced by European settlers) and American bison. They were among the largest flesh-eating land animals that lived during the Ice Age. Compared to modern African lions, they attained enormous size (25% larger) and had relatively long, slender limbs.
Panthera schaubi, currently classified as Puma schaubi, was a short-faced leopard-like cat about the size of a small leopard or large lynx; it is believed to be an Old World puma. Fossil leopards have been found in France and Italy, but in small quantities suggesting they were not prevalent in Europe. Felis lacustris ("Lake Cat"), also appears to be a North American Pliocene puma.
Panthera gombaszoegensis (P onca gombaszoegensis), the European Jaguar, was present around 1.6 million years ago and was larger than early American jaguars, probably hunting larger prey. They ranged across Italy, England, Germany, Spain, and France. Although currently given its own classification, P toscana, the Tuscany Lion (aka Tuscany Jaguar), may turn out to be a synonym for P gombaszoegensis. A form similar to P gombaszoegensis has been found dating from early Pleistocene East Africa and had both lion- and tiger-like characters.
"Pleistocene Tigers" have been found in Alaska, North America and are dated at only 100,000 years ago. There is debate over whether these were tigers or lions since the two species are structurally similar, resulting in some authorities giving the American tigers a separate classification. Pleistocene jaguars (approx 1.5 million years ago) were found as far north as Washington state and Nebraska, USA. Similar in size to modern jaguars they probably had similar lifestyles and pursued similar prey. The modern jaguar's range is South America and Central America, although some individuals have been found as far north as Texas and Florida, USA.
Lynx were known to be present in North America and Eurasia in Pliocene or Pleistocene time. The common ancestor of modern lynx and bobcats was probably a North American lynx of 6.7 million years ago. The extinct Issoire Lynx (Lynx issiodorensis) of 4 million years ago was larger than modern lynx with shorter legs. Modern lynx are smaller, a trend that is true in other species such as cheetah, jaguar, leopard and lion where the prehistoric forms were giants compared to their modern descendents. Their larger size was either an adaptation to colder climes or to cope with larger prey species.
A Pliocene Lynx from Britain was described as Felis (Lynx) brevirostris in 1889. Bobcats are related the Lynx. Earliest records of Bobcats (Lynx rufus, Felis rufus) date back 3.2 to 1.8 million years ago. It is the descendant of an earlier and larger Pleistocene species Lynx issiodorensis. A predominant Pleistocene subspecies of Bobcat was Lynx rufus calcaratus (Irvingtonian Bobcat), which was slightly larger than modern Bobcats. Another Pleistocene subspecies was Lynx rufus koakudsi.
North American Pliocene small cats include: Felis lacustris ("Lake Cat"), once considered to be a lynx, but now believed to be a puma; F rexroadensis which could be either a lynx or a leopard; F protolyncis ("Early Lynx") and F longignathus, which both resemble Lynx.
Genus Felis evolved around 12 million years ago. This genus Felis eventually gave rise to many of our small cats. They are known as "purring cats" because of the structure of their throat.
Two of the first modern Felis species were Felis lunensis (Martelli's cat, extinct), and Felis manul (Manul or Pallas's Cat). The ancestor of modern Felis species, including domestic cats, was F attica [syn F christoli].
Extinct Felis species are: F attica (primitive cat), F bituminosa, F daggetti, F issiodorensis, F lunensis and F vorohuensis (Pleistocene cat). The following debatable species may also be found in literature: F maniculus, F wenzensis, F antediluvia ("Cat from before the Biblical Flood"), F vireti, F (Sivafelis) obscura.
Fossils of a presumed prehistoric margay, Leopardus amnicola ("River Margay"), have been found in Arizona and Florida, USA.
We have a fairly good idea of the size and shape of prehistoric cats and cat-like creatures, but what did they actually look like? Did prehistoric "lions" they have manes and tail tufts? Did the sabre-tooth "tiger" have stripes? Where they sleek or shaggy? Prehistoric cats and cat analogues were shaped by the same forces that shaped modern cats, allowing us to create an image of what they looked like.
Like modern cats, most were ambush hunters, lying in wait for prey or stalking it before making a final dash. This means they needed camouflage. Spots and swirls break up the outline of a predator in undergrowth, while plain sandy or tawny hues blend into drier or barer backgrounds. The cats that hunted on open plains were probably sandy brown or tawny like modern cougars and lions, possibly with faint dappled markings. Others were probably more greyish in colour, like modern northern lynxes. Forest and woodland forms would need leopard-like or jaguar-like markings to blend in with dappled shade, though some may have been black. Although we speak of sabre-tooth "tigers", they would not have been striped unless they lived in habitats similar to those of modern tigers e.g. woodland with twiggy undergrowth or tall reeds . Depending on their habitat, the smaller cats would most likely have been similar to modern Ocelots (swirled), European and African Wildcats (faint mackerel tabby markings) or Leopard cats (spotted).
Those prehistoric big cats that lived in permanently snowy and icy climes would have needed paler coats to blend in with snow or patchy snow. They may have had coats resembling modern snow leopards. Some may have been almost white. They would need longer, denser fur to protect them from the cold, compared to those in warmer climes who would have shorter, sleeker coats Like modern cats, prehistoric cats in temperate zones (with summer and winter periods) would have moulted and grown longer coats for winter and shorter coats for summer. Modern male lions have manes, partly for social display and partly to protect their neck region during fights with competing males. There is some evidence that Smilodon was also social and lived in family groups, so Smilodon males may also have had manes or well-developed ruffs.
Most illustrations and reconstructions of prehistoric cats and cat analogues base their colour schemes on modern cats of comparable sizes living in comparable habitats. The remarkable similarity of cats living in similar habitats in different continents (e.g. lion and puma, jaguar and leopard) makes it reasonable to assume that ancient cats in corresponding habitats also evolved similar markings.
Older encyclopaedias show them as lions or tigers with long tails and over-sized teeth. The following image and texts are excerpts from "Wonders of Land and Sea" (?1913, edited by Graeme Williams). The text accompanying this depiction read "It is believed by some naturalists that the stripes of the sabre-toothed tiger ran horizontally along the body, while others maintain that they were as represented here." The image is a romanticised view of two Smilodons in the branches of a huge tree.
Before the Eocene was passing through the Oligocene into the Miocene, the carnivorous type had reached its most extravagant development, not merely in the ancestral Cat, but in the Sabre-toothed "Tigers." (A better name for this group is Machairodont - sabretoothed - as they were not specially allied to tigers; the tiger is a true cat of very modern origin.) The Machairodonts were the extreme of mammalian ferocity - felines preserving some primitive features lost by modern eats, but adapted in teeth and jaws for slaying beyond any other mammal.
The mouth of the Sabre-toothed Tiger (which type did not reach its culmination, however, till the close of the Miocene), could open with a gape that must have measured a foot and a half in the biggest examples. And thus the twelve-inch long, flattened, sabre-like, serrated canines of the upper jaw could be plunged for all their length into the neck or body of some great herbivore, aided by the leverage exercised by the prolonged chin, with its massed incisors and stunted canines. The molar teeth, reduced to two on each side, served only to sever, not to masticate, parcels of flesh. These gobbets must have been swallowed whole; or possibly the extreme form of the Machairodont lived mainly by blood-sucking, after severing the great arteries with its teeth. The biggest and most awful of the Sabre-tooths (Smilodon neogeus) lived in South America, lingering on almost to the human period in that region; while in Europe and Asia its near allies of the same genus were certainly contemporaneous with man. Indeed, in England the Sabre-toothed "Tiger" was possibly still in existence 100,000 years ago, when Paleolithic man had begun to take possession of the caves.
The True Cats, as distinct from the Machairodonts, apparently originated in Europe, and did not reach America till quite late in geological history. The Machairodonts were also European in their birthplace, finding their way to North America at a later date. But although America at the present day has no feline larger than the Jaguar, it seems to have possessed - at any rate in north and northernmost America, as late as the Pleistocene - cats as large as the Lion and Tiger; in fact, in all probability, near relations of the Tiger. The Tiger would seem to be of Asiatic origin, and never to have reached Europe, but to have extended its range to Arctic Asia (where its bones testify to its existence at no very distant date, well within the Arctic circle, on the New Siberian islands), and perhaps across the former Bering Isthmus into Arctic and Temperate North America. The Lion, which like the Jaguar and Tiger sprang from a Leopard ancestor (from a stock chiefly rosetted and spotted, but also striped), is most at home - so to speak - in France. The most primitive and jaguar-like type of Lion hitherto discovered has been found in early Pliocene strata in Southern France, while the extremist development of the Lion type also came into existence in France - the Cave Lion, which extended its range over nearly all Germany, England, North Italy, Switzerland, and Austria. Its somewhat degenerate descendant, the modern Lion (differing only from its ancestor by its much smaller size) ranged far afield, into Spain, Africa, Western Asia, and India.
The various sabre-tooth cats and cat-analogues were adapted to hunting the mega-fauna that inhabited various continents until the end of the last ice age. Changes in climate and habitat (and possibly other factors such as human hunters and major viral illnesses) led to the extinction of the mega-fauna. When prey species goes extinct, specialised predators such as sabre-tooth cats also become extinct. Those that could adapt to hunting smaller, more agile prey (e.g. hoofed grazing animals) went on to become modern cats. The population levels of elephants, our largest land mammals, are probably too small to sustain a population of sabre-toothed predators. There is no need for modern big cats to evolve into sabre-tooth forms; it would expend energy on growing the huge teeth but gain no competitive edge over other predators.
The modern Clouded Leopard is the closest we have to a sabre-tooth cat. It has the longest canines proportional to body size of any of the modern cats - the length of the fangs in approximately three times greater than the width of the fang at the socket. The backs of the canine teeth are very sharp, like those of the prehistoric sabre-toothed cats. There is a large gap between the canines and premolars, enabling them to take large bits out of their prey. Given a few million years (if humans don't wipe it out) in which to evolve and the right selective pressures, Genus Neofelis could give rise to the next generation of sabre-tooths.
There are cryptozoological reports of sabre-tooth cats surviving to the modern day in remote areas. The Tigre de Montage (Mountain Tiger) of northern Chad is described as lion-sized, striped reddish and white, tailless (or bobtailed) and having huge projecting fangs. From a selection of images, the one chosen by a Zagaoua hunter was Machairodus, an African sabre-tooth officially extinct for the last million years. The region is remote, mountainous and not well-known in zoological terms. Similar tales have come from the mountainous regions of Ecuador, Columbia, and Paraguay in South America, a region that has harboured marsupial sabre-tooths and eutherian sabre-tooth cats. In 1975, a big cat with 12 inch fangs was apparently killed in Paraguay; it was officially identified as a mutant Jaguar and unofficially identified as a Smilodon (the carcass seems to have been lost, preventing further study). Without firm evidence, e.g. a fresh or living specimen, sabre-tooth cats remain officially extinct.
Although they are often called "big cats" many of the prehistoric species were not true cats, but were cat-like mammals. South America and Australia were both island continents and lacked placental mammals. The "lions" and "tigers" of those continents were lion-like marsupials, more closely related to kangaroos, wombats and their like. Theyare known as "cat analogues" as they resembled cats and filled the same ecological niche as cats. Another line of prehistoric cats were the Paleofelids ("ancient cats") that developed in parallel with the true cats and from a common ancestor, but which have left no modern descendants. Finally there are the prehistoric true cats, extinct relatives of modern cats. Why would evolution create creatures very similar in form to cats? It's an example of convergent evolution - there are a limited number of solutions to environmental conditions hence animals that aren't closely related often independently evolve similar traits as they both adapt to similar environments and niches. The cat-like form has evolved at least three times: marsupial lions/tigers, Paleofelids and Neofelids. Sabre-toothed cat-like animals evolved separately four times in evolutionary history: Nimravids, Felids, Creodonts and the Thylacosmilids.
The taxonomy (classification) of both living and extinct species changes frequently. As more fossil evidence comes to light, species are reclassified. Some are given their own species or genus while others are absorbed into an existing species or genus and their original classification is scrapped.
Although there are references to animals being found in places far apart on the modern world map, the continents used to look very different. Some land masses that were once joined together have now split and drifted apart, others that were far apart have collided. Some land masses that are currently not joined to each other were joined by ancient land bridges when sea levels were much lower than they are today.
Carnivorous mammals evolved from Miacids small pine marten-like insectivores that lived 60 million - 55 million years ago. The miacids split into two lines: Miacidae and Viverravidae. Miacidae gave rise to Arctoidea/Canoidea group (bears and dogs) while Viverravidae gave rise to Aeluroidea/Feloidea group (cats, hyenas, civets, mongooses) around 48 million years ago. The Viverravidae also gave rise to a group called Nimravidae. The Nimravids were cat-like creatures that evolved in parallel with true cats; they are not part of true cat lineage and have left no living descendents.The first true cat to arise from Viverravidae was Proailurus (first cat") around 30 million years ago. The best-known species was P lemanensis, found in France.Proailurus was a small weasel-like cat with relatively short legs and a long body. It had one more premolar on each side of its bottom jaw than do modern cats. About 20 million years ago, Proailurus gave rise to Pseudaelurus. Pseudaelurus were Miocene ancestors of cats. Pseudaelurus lorteti was about the size of a large lynx while P validus was the size of a large lynx or small puma. Three other species of early cat are described as Pratifelis, Vishnufelis and Sivaelurus (S chinjienis).Pseudaelurus went on to split into two major groups: the Machairodontinae (true sabre-tooths) and Schizailurus (the ancestor of the modern day Felidae group).
18 million years ago, Schizailurus gave rise to the Felidae. The first of the modern Felids were the early cheetahs; now represented by Acinonyx (modern cheetah); true cheetahs are believed to have evolved around 7 million years ago. Some sources claim Miracinonyx (North American cheetahs) evolved only 4 million years ago from Acinonyx, but recent studies show Miracinonyx was probably ancestral to both cheetahs and puma and was intermediate in type between these two modern species.
Around 12 million years ago, genus Felis appeared and eventually gave rise to many of our small cats. Two of the first modern Felis species were Felis lunensis (Martelli's cat, extinct), and Felis manul (Manul or Pallas's Cat, living). Extinct Felis species are: F attica, F bituminosa, F daggetti, F issiodorensis (Issoire Lynx), F lunensis and F vorohuensis. The ancestor of modern Felis species was F attica. Genus Panthera ("biting cats" or "roaring cats") genera evolved around 3 million years ago; there are a number of extinct species discussed later in this article.
Genera Acinonyx, Felis and Panthera are all represented today and taxa of some modern species is regularly revised as more complete fossils of ancestral species are found, giving a clearer indication of who begat whom and when various lineages split.
The jaguar-sized Thylacosmilus ("pouched blade") was a large, predatory marsupial; part of a unique group of predators on the South American pampas; the borhyaenids. These appeared in the Miocene and ruled the South American forests and woodlands for some 30 million years, but have no known ancestor or descendant. Thylacosmilus was the most successful member of that family and was the ultimate mammalian predator of its day in Plio-Pleistocene South America, but when the continents of North America and South America joined, it lost out to the more highly developed and faster eutherian cats.
Two species are described: T atrox and T lentis. Thylacosmilus atrox looked like a sabre-toothed cat, but is more closely related to kangaroos. As far as we know, Thylacosmilus was the only marsupial to have developed the sabre-toothed weapon. Like Smilodon, the eutherian sabre-tooth tiger, it had adapted to hunting mega-fauna.
Thylacosmilus lacked incisor teeth, but had very long upper canine teeth that grew continually. These long stabbing teeth projected below the mouth-line. Strong neck and jaw muscles allowed the sabre-teeth to be driven downward with a tremendous killing force. Its huge stabbing teeth were about 15 cm (6 inches) long (longer than those of Smilodon) and may have been used to slash the soft throat of its prey. The jaws were capable of a gape that left the teeth clear to do their work. These sabres grew continually throughout Thylacosmilus's life, much like the incisors of modern rodents. Unlike Smilodon (see later), it had no scabbard-like tooth-guards on its lower jaw though its skull had a deep flange on its lower jaw, forming a protective sheath for when the sabre teeth were not being used.
Unlike modern cats, which tend to be sleek and long-legged, it appears to have been short-legged and heavily built, being about 1.2 metres (4 ft) long and weighing around 100 kilos. Its claws were not retractile. It probably preyed on large, slow-moving mammals and when the two continents joined, the highly specialised Thylacosmilus could not compete against the faster, sleeker eutherian big cats. South America has also had at least three species of cats whose body weights exceeded 300 kilos - about twice the weight of modern lions.
The Thylacoleonidae were lion-like marsupials that inhabited Australia in Oligocene to Pleistocene times. They probably hunted across the Australian grasslands, although some may have been arboreal. They were vombatomorphian (wombat-like) marsupials, evolved from herbivore ancestors; their closest living relatives being koalas and wombats. The more primitive species had generalised crushing molar teeth (like modern omnivores) as well as carnassial blades. In more specialised species, the crushing molars were reduced or absent and the carnassials had become huge.
The Thylacoleonidae ranged from the size of a domestic cat to the size of a leopard and possibly even the size of a lion (1.7 metres/5 ft 6 in). So far, eight species of marsupial lion have been discovered and there may be at least two more. Those of genus Wakaleo were leopard-sized and designed for power rather than speed. W alcootaensis was slightly larger than W oldfieldi or W vanderleueri. These "marsupial leopards" may have ambushed prey from tree branches. Priscaleo was much smaller. P pitikantensis was about the size of a modern Australian possum. P roskellyae was about the size of a domestic cat, possibly up to ocelot-sized, and may have been arboreal.
The most famous member of this family is Thylacoleo carnifex, the "marsupial lion". This was Australia's equivalent to the South American marsupial Thylacosmilus atrox and to the eutherian Smilodon. Its enormous meat-shearing carnassial (cheek) teeth were the largest of any mammalian predator. It also had bolt-cutter incisors, and switch-blade-like claws on its semi-opposable thumbs. It was the most specialised mammalian carnivore ever known; entirely lacking grinding teeth. Because T carnifex lacked large canines, it was originally believed to be a herbivore, using its unusual front teeth and claws to break open nuts and fruit; its lack of grinding teeth suggest a diet of soft fruit such as melons! However, wear on the teeth indicates a meat-eating diet, and it probably preyed on giant kangaroos and wombats of the time. Compared to sabre-tigers, such as Smilodon, it had a short cat-like face and more elaborate carnassial teeth, giving it a powerful killing bite. Most modern cats have carnassials that can crunch smaller bones, but Thylacoleo's teeth lacked bone-crunching adaptations and were entirely adapted to shearing soft tissue. Projecting front incisors were modified into killing teeth, and looked rather like the canines in the placental carnivores; the actual canine teeth were insignificant.
T carnifex had a short body, closer in length to that of a leopard rather than a lion, but the bones of its legs show it was far more robust than a leopard. Estimates derived from size (partly based on skull size) and robustness suggest it weighed between 100 and 130 kilos, putting it in the same size range as modern tigers and lions. It was extremely robust and built for power rather than endurance, with tremendously powerful forelimbs. It probably ambushed prey as large as, or larger than, itself, using the thumb claws to hold the prey in a deadly embrace while employing its fang-like incisors.
T carnifex survived until around 50,000 years ago and may have come into conflict with early Aboriginal settlers entering Australia. Conflict and competition with humans, and with the introduced dingo, may have contributed to the extinction of this highly specialised carnivore. There are theories that relict populations of smaller marsupial lions survive in the form of the cryptozoological "Queensland Tiger". Like Thylacoleo, the Queensland Tiger is described as short-headed, sharp-clawed and superficially cat like. Eyewitnesses (and a single photograph) show it to have vertical striped on the forequarters. It has never been positively identified.
The Nimravidae were a separate family of cat-like animals that evolved parallel to the true cats (Felidae). The common ancestor of the Nimravidae and the Felidae was the Viverravidae (feline-like) group of miacids some 55 million years ago in the late Eocene. Proailurus, a descendent of the miacids, evolved into Pseudaelurus, which split into two main groups, one of which became the modern cats (Neofelids), and one became the Machairodonts (true sabre-toothed cats, a sub-family of the Felidae). Nimravids are also known as the Paleofelids (ancient cats), or False Sabre-toothed Cats (because they are cat-like, but not true cats). They were the earliest cats to evolve and lived from the late Eocene (36 million years ago) to the late Miocene (5 million years ago), peaking around 28 million years ago. The three subfamilies of nimravid and their genera are shown in the table below. Some are known from single or fragmented specimens and the taxonomy may change as more fossils are discovered.
Nimravids had different skulls to true cats. The structure of their middle and inner ear was different and many Nimravids had a flange on the front of the lower jaw (also seen in some machairodonts [true sabre-tooths] and thylacosmilids [marsupial sabre-tooth]). The flange is a bony prominence that projects downwards and is as long as the canine teeth; the teeth fit into a groove. Barbourofelis has the most prominent flange, while Nimravus and Dinaelurus lacked a flange. The actual sabres were narrow, pointed canines whose length varied according to species.
Nimravids were very cat-like in appearance and had retractile claws. Many were muscular and low slung with heavy-set bodies on short legs. Like the modern lynx, some were short-tailed although many others were long-bodied and long-tailed. The skulls of genera Dinictis, Nimravus and Dinaelurus are especially cat-like. Their prominent upper canines were longer than those of modern cats, but shorter than those of the true sabre-toothed cats; their lower canines were proportionally longer. The most common species in the fossil record are those belonging to Dinictis, Eusmilus, Hoplophoneus and Barbourofelis.
Hoplophoneus lived during the late Oligocene (33 - 30.5 million years ago), some 20 million years before Smilodon. Some earlier authors erroneously place Hoplophoneus among the Felidae (true cats) as the ancestor of Smilodon and true sabre-tooths, but current fossil evidence makes this incorrect and indicates that Hoplophoneus and Smilodon are from different evolutionary lines. Some were the size of bobcats while others were jaguar-sized. Like many other Nimravids, Hoplophoneous had a bony flange into which its curved canines fit.
Nimravus has been found in France and parts of North America from the early Oligocene to early Miocene. Some were 1.2 metres (4 ft) long. With its sleek body, it may have resembled the modern caracal, although it had a longer back and more dog-like feet with partially retractile claws. It competed with other false sabre-tooths such as Eusmilus. A Nimravus skull, found in North America, had been pierced in the forehead region, the hole exactly matching the dimensions of Eusrnilus' sabre tooth; Nimravus survived as the wound showed signs of healing. It probably hunted birds and small mammals, ambushing them like modern cats, rather than chasing them down. Some specimens still have unclear classification, for example a species once identified as N catacopis is closer to true cats and is now classified as Machairodus aphanistus (previously Machairodus catacopis).
Dinictis was a small nimravid that lived on the plains of North America during the late Eocene and early Oligocene (40 million years ago). Dinictids had a sleek bodies, short legs, long tails, and walked plantigrade ("on the whole foot" modern cats walk digitigrade "on the toes").
Eusmilus was a dirk-toothed cat found in France and parts of North America during the late Oligocene (30.5-28.5 million years ago). It was noted for its long, flattened sabres and very prominent mandibular flange. Most were leopard-sized and rather long-bodied and short-legged compared to modern leopards. Some reached 2.5 metres (8 ft) long. It was a typical false sabre-tooth with enlarged upper canines, but insignificant lower canines, while many of the other teeth had been lost to accommodate its sabres (Eusmilus had 26 teeth, compared to 44 teeth in other carnivores). The jaw hinge was modified to open to an angle of 90 degrees to allow the great sabre teeth to do their work. Its lower jaw had bony guards that lay along the length of the sabres, protecting them from damage when the mouth was closed. There is fossil evidence of conflict between Eusmilus and Nimravus.
The lion-sized Barbourofelis was one of the most recent members of the Nimravidae family. It lived during the late Miocene (15 million - 6 million years ago) and had the longest canines of all the nimravids. They had very prominent flanges on the lower jaws and an unusually shaped skull. The Barbourofelids were probably very muscular, resembling a bear-like lion or lion-like bear. Pogonodon was a cat-like sabre-tooth.
The Machairodontinae are true cats and their fossils have been found in North America, Europe, Asia and Africa. Although we tend to think of the sabre-toothed tiger, there were two varieties of sabre-toothed cats: dirk-toothed cats and scimitar-toothed cats. Dirk-toothed cats had two long, narrow upper canines, and were usually short-legged and stocky. Scimitar-toothed cats had upper canines that were shorter and broader, longer, thinner legs and were generally more lithe. The exception was a cat known as Xenosmilus, which has the short, broad canines of a scimitar-toothed cat, but has short legs.
Modern cats have conical canine teeth, but the machairodonts' (machairodont means "sabre tooth") canines were flattened from side to side (like a blade) as well as being elongated. To accommodate their large canines, they had fewer upper premolar teeth. Their incisor teeth were larger, angled differently and placed further forward than in modern cats. Other adaptations allowed them to open their jaws extremely wide and gave them strong neck muscles. Some species, such as Megantereon, had a bony flange that protruded downward from the front of their lower jaw.
Although the sabre-toothed cats have long dagger-like canines, these were probably too blunt and fragile to be used to stab prey. They were unlikely to have gone for the nape of the neck to sever the spine, like many modern cats. If they hit bone, they could shatter (leading to abscesses and possibly fatal bacterial infections). The current theory is that sabre-toothed cats went for the soft throat of their prey, using their powerful teeth to sever the arteries and windpipe.
These were Eurasian scimitar-toothed cats that lived during the late Miocene to early Pleistocene.
The best known genus is Dinofelis ("giant cat") which lived in Eurasia, Africa and North America around 5 million - 1.5 million years ago. In South Africa, Dinofelis has also been found at sites along with Australopithecines, so it was probably a predator of our own ancestors. Genus Dinofelis includes cats previously classified as Therailurus. It became extinct in Eurasia and North America during the Early Pleistocene, but survived in Africa until the Mid-Pleistocene. The largest known form was the Chinese D abeli. Its size and build are comparable to a large leopard or jaguar (1.2 metres/4 ft) with forelimbs more heavily built than hind-limbs. Like the modern jaguar, they may have been forest-dwellers. Dinofelis ("giant cat") had flattened canines considerably shorter than those of the sabre-tooths, scimitar-tooths or even dirk-tooth cats. The canines were longer than those of biting cats (those that kill prey with a single neck-bite) so it is debatable as to which subfamily of the Dinofelis belongs.
Metailurus lived 15 - 8 million years ago and were similar to Dinofelis in body type and shape. They were smaller than Dinofelis, being leopard sized, but had longer canine teeth, intermediate between modern cats and sabre-toothed cats. Like Dinofelis, Metailurus was stockily built with forelimbs more heavily built that its hind limbs. Its build suggests it hunted in forested areas and may have been more arboreal than Dinofelis.
The puma-sized Adelphailurus is known from a single type specimen from Kansas, USA. Unusually for a cat, Adelphailurus kansensis retained the upper second premolar, a trait it shared with Stenailurus. The members of genus Pontosmilus were formerly classified as Paramachairodus.
This comprises the genera Machairodus, Homotherium and Xenosmilus. Classification is liable to be revised as more fossils are discovered and there are museum specimens assigned to 4 other genera.
Machairodus is a variable genus of large sabre-toothed cats ranging in size and structure from smaller varieties right up to lion-sized. They were found in Europe, Asia, Africa and North America from 15 - 2 million years ago. The taxonomy is debatable. They have large canine teeth, and its incisors, canines and carnassials has serrated edges. There were two basic types of Machairodus: the primitive type and the evolved type (possibly adaptations to different environments). The more primitive types included M aphanistus and resembled Smilodon. The more evolved type had serrated teeth and elongated forelimbs structurally similar to the hyena-like Homotherium; they may have been ancestral to Homotherium. The variable forms indicate a adaptations to different environments ranging from forest/woodland dwelling to plains hunting. The hyena-like species may have covered long distances while hunting or for opportunistic scavenging.
Homotherium (therium = "beast") is a group of unique hyena-like sabre-toothed cats that also ranged widely (Africa, Asia, Europe, North America) from 3 million - 0.5 million years ago. They were about 1.2 metres (4 ft) long with front limbs longer than the rear ones. Homotherium's incisors were very large and robust and they had serrated medium-length canine teeth. Homotherium is a scimitar-toothed cat i.e. it has shorter, flatter canines than other sabre-tooth cats and its canines curve backwards like scimitar blades. Homotherium would have had the sloping look of a hyena with slender legs and relatively long neck. Its anatomy suggest that it walked with the whole foot on the ground (plantigrade) like a bear. This hyena-like conformation may have allowed them to cover long distances when hunting. It is more likely to have walked semi-plantigrade, the back sloping slightly; an adaptation for greater strength.
Hometherium survived until the end of the last ice age about 14,000 years ago and probably preyed on mammoths, possibly hunting in family groups. In Texas, the bones of a family group of scimitar-tooths are preserved alongside young mammoths and their eventual extinction was probably linked to a decline in prey species. As an adaptation to ice age conditions, some species may have been white or pale grey (like modern arctic predators).
H serum, the North American scimitar cat ,was originally named Dinobastis serus. It was short-tailed and slender-limbed, with relatively long forelimbs and short, powerful hindlimbs. Its deepened chin meant that its upper canines did not protrude beyond the lower margin of the lower jaw. H serum's large nasal opening, like that of the cheetah, would have allowed quicker oxygen intake aiding in rapid running. Skulls show it had a large visual cortex, indicative of a daytime hunter. It was built for short bursts of speed, rather than long chases. The claws of its forelimbs were not retractile, allowing better traction at high speed. Its hind limbs were shorter than its forelimbs and had a bear-like heel and ankle. The long hind feet had non-retractile claws.
H latidens is depicted in paleolithic stone carvings from Isturitz, south-western France show a short-tailed big cat with a deeply set lower jaw. This matches the traits of the European Scimitar-tooth, H latidens. The carving suggests that the cats had spotted pelts and paler undersides. H ultimum, the Asian scimitar cat, occurred in China.
Homotherium ischyros (or Ischyrosmilus), had canines serrated like steak-knives, along their front and back edges. This made it easier to slice through the skin of thick-skinned prey. Megantereon lacked these serrations on its upper canines. Ischyrosmilus's exact taxonomy is unclear and it may be one of the Smilodontini.
Xenosmilus ("strange knife") lived in the Florida, USA region 1.7 - 1 million years ago and was a robust cat with short legs and very thick, stout canine teeth. It was lion-sized, very robust and somewhat bear-like Pleistocene felid (length 2 metres / 7 ft, weight 180-230 kg / 400-500 lbs). Like Homotherium, it had broad, knifelike, coarsely crenulated teeth and projecting long, curved, serrated incisors, but it had the short, stout-legged features of Smilodon. Originally palaeontologists thought they had mixed up the bones of 2 other species since dirk-tooth cats were bear-like with two long, narrow upper canine teeth and short legs, while scimitar-tooth cats were longer-legged with two shorter, broader upper canines. Xenosmilus had a mix of features: the short, broad coarsely crenulated teeth of a chasing cat, but the stocky legs of an ambush hunter. It probably stalked close to its prey and then sprinted from cover to catch it. The specimens were found with bones of peccaries (wild pigs), giving an indication of its main prey. It appears to have been a more specialised sabre-toothed cat than Smilodon and its size made it the most ferocious sabre-tooth in the world at the time. Smilodon may only have become a dominant predator after Xenosmilus vanished.
This contains the famous sabre-toothed "tiger", Smilodon ("blade tooth"), of the late Pleistocene age (1.5 million - 10,000 years ago). The three known species were widespread throughout the Americas. Smilodon was stocky, with short, powerful legs and a bobtail. Their canine teeth were the longest of all the true sabre-toothed cats, being about 18 cm long. The South America Smilodon populator was the largest species. The South American Smilodon neogaeus is probably a synonym for S populator. The more famous Smilodon fatalis was found across North and South America, having migrated there from North America during the Pleistocene. S fatalis is sometimes divided into two separate species: S californicus and S floridanus (these may have been sub-species or variant populations i.e. S fatalis californicus and S fatalis floridanus). They are usually compared to the modern lion based on body length (1.2 metres/4 ft) and conformation, but were more robustly and powerfully built. S fatalis and S populator were around 11/2 - 2 times heavier than the average lion (around 170 kilos). Smilodon gracilis was the smallest of the Smilodons and the earliest species (about 2.5 million years ago); it was found in the eastern United States and weighed around 80 kilos.
Smilodon's anatomy shows them to be were specialised hunters of big game; they probably ambushed their prey - their robust build is not designed for chasing it down. Most modern cats have carnassials that can crunch smaller bones, but Smilodon's teeth lacked bone-crunching adaptations and were entirely adapted to shearing soft tissue. The muscles of its shoulders and neck were arranged to produce a powerful downward lunge of its massive head. The jaw opened to an angle of over 120 degrees, to allow the huge upper canines to be driven into prey. The canines were oval in cross-section to retain strength, but also to ensure minimum resistance as they were sunk into the prey. They were also serrated along their rear edges, so they pierced flesh more easily. They probably preyed on large, slow-moving, thick-skinned herbivores, but also scavenged dead and dying animals. More than 2000 Smilodon skeletons have been recovered from the Pleistocene tar pits of La Brea (Los Angeles, USA) where they had been fatally lured by large animals trapped in the tar.
It seems likely that Smilodon lived in family groups, much like modern lions, and possibly hunted in groups. Some specimens showed signs of healed fractures, suggesting they ate food caught by family members. Their extinction seems to have coincided with open plains taking over from forest/woodland. Smilodon was not built for the chase and this reduction in cover would have made it harder to ambush prey.
Megantereon was another genus of cats with impressive canine teeth, although they are not well-represented. About the size of a jaguar (1.2 metres/4 ft), they flourished in the Mediterranean and spread throughout Africa, Eurasia and North America around 3 million - 1 million years ago. The only complete skeleton was found in France. They had very large upper canines but relatively small lower ones. Though impressive, its teeth were more like daggers than sabres in size and shape, hence Megantereon and its immediate relatives are referred to as dirk-tooth (dagger-tooth) cats. They are also characterized by their prominent mandibular flange. Only a single species, Megantereon cultridens, is known from fossil records, although some Chinese specimens are known as M. nihowanensis. They are believed by most to be the direct ancestors of Smilodon and other later sabre-tooth cats.
Also in this group is Paramachairodus, although it is under much debate as to the placement of this genus. Many animals formerly placed in this genus have been reassigned to Pontosmilus and placed within Metailurini. They are thought to have existed between 20 - 9 million years ago. The two species known are Paramachairodus ogygia (Spain) and Paramachairodus orientalis, plus the disputed P maximiliani..
Modern Felids evolved around 18 million years ago. The first of these were the early cheetahs; now represented by Acinonyx (modern cheetah); true cheetahs are believed to have evolved around 7 million years ago. Some sources claim Miracinonyx (North American cheetahs) evolved only 4 million years ago from Acinonyx, but recent studies show ("miracle cheetah") was probably ancestral to both cheetahs and puma and was intermediate in type between these two modern species. Another cheetah was the Sivapanthera genus.
Cheetah-like cats arose around 18 million years ago. According to some studies, the ancestor of modern cheetahs originated in Africa during the Miocene and later migrated, giving rise to the now-extinct North American cheetahs. More recent studies suggest that a North American cheetah called Miracinonyx was the ancestor of both African cheetahs (modern Acinonyx) and American pumas (Puma concolor). Miracinonyx would have migrated across continents during the Ice Age. Miracinonyx inexpectatus [M studeri] existed in North America during the early Pleistocene (1 to 1.5 millions years ago) and may be even older. It had proportions intermediate between the modern cheetah and modern pumaAs a result, iIt is sometimes linked to the cryptozoological Onza (a gracile form of puma). Two species of cheetah inhabited late Pleistocene North America (100,000 years ago): Miracinonyx inexpectatus [M studeri] and Miracinonyx trumani.
Fossil evidence of early cheetahs is fragmentary, but Miracinonyx resembles modern cheetahs in having a short face, wide nasal passages and long body, but were less lanky. M inexpectatus and M trumani may be the reason North American evolved into such fast sprinters; North America has no living predator able to match the pronghorn's in speed. Unlike modern cheetahs, Miracinonyx inexpectatus had fully retractile claws and more robust conformation with shorter limbs than modern cheetahs. M inexpectatus would have been faster than the puma, but not as accomplished a sprinter as modern cheetahs; it was also better equipped for climbing.
The early true cheetah, Acinonyx pardinensis, appeared during the Pliocene and at 200 lbs were much larger than modern cheetahs. Known as Giant Cheetahs, they became widespread in China, southern Europe and India throughout the Ice Age, were lion-sized cheetah and probably as fast as modern cheetahs. Intermediate-sized cheetahs, Acinonyx intermedius, ranged from Africa as Far East as China during the mid-Pleistocene and became adapted to hunting on open grassland. These were larger than modern cheetahs. Acinonyx parchidinensis was the Pleistocene cheetah. The smaller modern cheetah, A jubatus, was once much more widespread, but became extinct in eastern Asia at the end of the Ice Age.
Biting cats are so-called because their relatively short, strong canines are adapted to dispatching prey by biting the bones and sinew of the neck and throttling it. Genus Panthera genera evolved around 3 million years ago and these have become the modern day big cats ("roaring cats" or "biting cats"). Prehistoric relatives of modern big cats lived between the Pleistocene to Recent times and ranged across South Africa, Asia, Europe and North America. Some, such as the "cave lion" were truly impressive creatures, reaching 3.5 metres (111/2 ft). The modern lion, Panthera leo, is now restricted to parts of Africa and to western India. There are extinct sub-species of the modern lion; until recent times there were sub-species in Arabia and Iran (the Barbary lion has been rediscovered and is being conserved).
Primitive lions (P leo fossilis) dispersed in the Old World about 500,000 years ago. P youngi, from north-eastern China 350,000 years ago, had similarities to both P leo spelaea (European Cave Lion) and P leo atrox (North American Cave Lion) and may be a link between the Eurasian/American cave lions and Asian/African modern lions. Extinct Panthera species include: P atrox (P leo atrox) (American lion), P gombaszoegensis (European jaguar), P leo spelaea (European Cave Lion), P palaeosinensis (a primitive leopard), P pardoides (a primitive leopard), P schaubi (a short-faced leopard) and P toscana (Tuscany lion, Tuscany jaguar). Some of these classifications are being revised as more complete specimens are discovered. Panthera schaubi (syn Viretailurus schaubi), which resembles a short-faced leopard, is currently regarded as an Old World puma, Puma schaubi.
Panthera leo spelaea, the European Cave Lion (300,000 to 10,000 years ago), was probably the largest cat that ever lived, being around 25% larger than modern lions, and larger than modern Siberian tigers. It was probably comparable in size to modern liger hybrids. It ranged across most of Europe and is depicted in cave paintings. Cave paintings and remains show that it lived until historical times in south-eastern Europe and possibly as recently as 2000 years ago in the Balkans. Cave paintings from Germany show cave lions as having ruffs or manes and tufted tails. A wall engraving from France and an ivory figurine of a lion-pelt-wearing human from Russia indicate faintly striped pelts. European Cave Lions inhabited steppes and parkland regions in the north and semi-desert areas in the south of Eurasia. They were evidently not adapted to deep snow or to dense forests.
Panthera atrox (P leo atrox) was a North American lion whose range extended to northern South America (Peru). P atrox crossed to North America over the Bering Strait land bridge during the last ice age, about 35,000 to 20,000 years ago. Its remains have been found in Alaska and some specimens have been found in the La Brea tar pits, Los Angeles, USA. Relatively few P atrox fossils have been recovered from La Brea compared to fossils of other carnivores; this lion may have been intelligent enough to avoid the natural traps. It probably hunted deer, North American horses (which became extinct and were reintroduced by European settlers) and American bison. They were among the largest flesh-eating land animals that lived during the Ice Age. Compared to modern African lions, they attained enormous size (25% larger) and had relatively long, slender limbs.
Panthera schaubi, currently classified as Puma schaubi, was a short-faced leopard-like cat about the size of a small leopard or large lynx; it is believed to be an Old World puma. Fossil leopards have been found in France and Italy, but in small quantities suggesting they were not prevalent in Europe. Felis lacustris ("Lake Cat"), also appears to be a North American Pliocene puma.
Panthera gombaszoegensis (P onca gombaszoegensis), the European Jaguar, was present around 1.6 million years ago and was larger than early American jaguars, probably hunting larger prey. They ranged across Italy, England, Germany, Spain, and France. Although currently given its own classification, P toscana, the Tuscany Lion (aka Tuscany Jaguar), may turn out to be a synonym for P gombaszoegensis. A form similar to P gombaszoegensis has been found dating from early Pleistocene East Africa and had both lion- and tiger-like characters.
"Pleistocene Tigers" have been found in Alaska, North America and are dated at only 100,000 years ago. There is debate over whether these were tigers or lions since the two species are structurally similar, resulting in some authorities giving the American tigers a separate classification. Pleistocene jaguars (approx 1.5 million years ago) were found as far north as Washington state and Nebraska, USA. Similar in size to modern jaguars they probably had similar lifestyles and pursued similar prey. The modern jaguar's range is South America and Central America, although some individuals have been found as far north as Texas and Florida, USA.
Lynx were known to be present in North America and Eurasia in Pliocene or Pleistocene time. The common ancestor of modern lynx and bobcats was probably a North American lynx of 6.7 million years ago. The extinct Issoire Lynx (Lynx issiodorensis) of 4 million years ago was larger than modern lynx with shorter legs. Modern lynx are smaller, a trend that is true in other species such as cheetah, jaguar, leopard and lion where the prehistoric forms were giants compared to their modern descendents. Their larger size was either an adaptation to colder climes or to cope with larger prey species.
A Pliocene Lynx from Britain was described as Felis (Lynx) brevirostris in 1889. Bobcats are related the Lynx. Earliest records of Bobcats (Lynx rufus, Felis rufus) date back 3.2 to 1.8 million years ago. It is the descendant of an earlier and larger Pleistocene species Lynx issiodorensis. A predominant Pleistocene subspecies of Bobcat was Lynx rufus calcaratus (Irvingtonian Bobcat), which was slightly larger than modern Bobcats. Another Pleistocene subspecies was Lynx rufus koakudsi.
North American Pliocene small cats include: Felis lacustris ("Lake Cat"), once considered to be a lynx, but now believed to be a puma; F rexroadensis which could be either a lynx or a leopard; F protolyncis ("Early Lynx") and F longignathus, which both resemble Lynx.
Genus Felis evolved around 12 million years ago. This genus Felis eventually gave rise to many of our small cats. They are known as "purring cats" because of the structure of their throat.
Two of the first modern Felis species were Felis lunensis (Martelli's cat, extinct), and Felis manul (Manul or Pallas's Cat). The ancestor of modern Felis species, including domestic cats, was F attica [syn F christoli].
Extinct Felis species are: F attica (primitive cat), F bituminosa, F daggetti, F issiodorensis, F lunensis and F vorohuensis (Pleistocene cat). The following debatable species may also be found in literature: F maniculus, F wenzensis, F antediluvia ("Cat from before the Biblical Flood"), F vireti, F (Sivafelis) obscura.
Fossils of a presumed prehistoric margay, Leopardus amnicola ("River Margay"), have been found in Arizona and Florida, USA.
We have a fairly good idea of the size and shape of prehistoric cats and cat-like creatures, but what did they actually look like? Did prehistoric "lions" they have manes and tail tufts? Did the sabre-tooth "tiger" have stripes? Where they sleek or shaggy? Prehistoric cats and cat analogues were shaped by the same forces that shaped modern cats, allowing us to create an image of what they looked like.
Like modern cats, most were ambush hunters, lying in wait for prey or stalking it before making a final dash. This means they needed camouflage. Spots and swirls break up the outline of a predator in undergrowth, while plain sandy or tawny hues blend into drier or barer backgrounds. The cats that hunted on open plains were probably sandy brown or tawny like modern cougars and lions, possibly with faint dappled markings. Others were probably more greyish in colour, like modern northern lynxes. Forest and woodland forms would need leopard-like or jaguar-like markings to blend in with dappled shade, though some may have been black. Although we speak of sabre-tooth "tigers", they would not have been striped unless they lived in habitats similar to those of modern tigers e.g. woodland with twiggy undergrowth or tall reeds . Depending on their habitat, the smaller cats would most likely have been similar to modern Ocelots (swirled), European and African Wildcats (faint mackerel tabby markings) or Leopard cats (spotted).
Those prehistoric big cats that lived in permanently snowy and icy climes would have needed paler coats to blend in with snow or patchy snow. They may have had coats resembling modern snow leopards. Some may have been almost white. They would need longer, denser fur to protect them from the cold, compared to those in warmer climes who would have shorter, sleeker coats Like modern cats, prehistoric cats in temperate zones (with summer and winter periods) would have moulted and grown longer coats for winter and shorter coats for summer. Modern male lions have manes, partly for social display and partly to protect their neck region during fights with competing males. There is some evidence that Smilodon was also social and lived in family groups, so Smilodon males may also have had manes or well-developed ruffs.
Most illustrations and reconstructions of prehistoric cats and cat analogues base their colour schemes on modern cats of comparable sizes living in comparable habitats. The remarkable similarity of cats living in similar habitats in different continents (e.g. lion and puma, jaguar and leopard) makes it reasonable to assume that ancient cats in corresponding habitats also evolved similar markings.
Older encyclopaedias show them as lions or tigers with long tails and over-sized teeth. The following image and texts are excerpts from "Wonders of Land and Sea" (?1913, edited by Graeme Williams). The text accompanying this depiction read "It is believed by some naturalists that the stripes of the sabre-toothed tiger ran horizontally along the body, while others maintain that they were as represented here." The image is a romanticised view of two Smilodons in the branches of a huge tree.
Before the Eocene was passing through the Oligocene into the Miocene, the carnivorous type had reached its most extravagant development, not merely in the ancestral Cat, but in the Sabre-toothed "Tigers." (A better name for this group is Machairodont - sabretoothed - as they were not specially allied to tigers; the tiger is a true cat of very modern origin.) The Machairodonts were the extreme of mammalian ferocity - felines preserving some primitive features lost by modern eats, but adapted in teeth and jaws for slaying beyond any other mammal.
The mouth of the Sabre-toothed Tiger (which type did not reach its culmination, however, till the close of the Miocene), could open with a gape that must have measured a foot and a half in the biggest examples. And thus the twelve-inch long, flattened, sabre-like, serrated canines of the upper jaw could be plunged for all their length into the neck or body of some great herbivore, aided by the leverage exercised by the prolonged chin, with its massed incisors and stunted canines. The molar teeth, reduced to two on each side, served only to sever, not to masticate, parcels of flesh. These gobbets must have been swallowed whole; or possibly the extreme form of the Machairodont lived mainly by blood-sucking, after severing the great arteries with its teeth. The biggest and most awful of the Sabre-tooths (Smilodon neogeus) lived in South America, lingering on almost to the human period in that region; while in Europe and Asia its near allies of the same genus were certainly contemporaneous with man. Indeed, in England the Sabre-toothed "Tiger" was possibly still in existence 100,000 years ago, when Paleolithic man had begun to take possession of the caves.
The True Cats, as distinct from the Machairodonts, apparently originated in Europe, and did not reach America till quite late in geological history. The Machairodonts were also European in their birthplace, finding their way to North America at a later date. But although America at the present day has no feline larger than the Jaguar, it seems to have possessed - at any rate in north and northernmost America, as late as the Pleistocene - cats as large as the Lion and Tiger; in fact, in all probability, near relations of the Tiger. The Tiger would seem to be of Asiatic origin, and never to have reached Europe, but to have extended its range to Arctic Asia (where its bones testify to its existence at no very distant date, well within the Arctic circle, on the New Siberian islands), and perhaps across the former Bering Isthmus into Arctic and Temperate North America. The Lion, which like the Jaguar and Tiger sprang from a Leopard ancestor (from a stock chiefly rosetted and spotted, but also striped), is most at home - so to speak - in France. The most primitive and jaguar-like type of Lion hitherto discovered has been found in early Pliocene strata in Southern France, while the extremist development of the Lion type also came into existence in France - the Cave Lion, which extended its range over nearly all Germany, England, North Italy, Switzerland, and Austria. Its somewhat degenerate descendant, the modern Lion (differing only from its ancestor by its much smaller size) ranged far afield, into Spain, Africa, Western Asia, and India.
The various sabre-tooth cats and cat-analogues were adapted to hunting the mega-fauna that inhabited various continents until the end of the last ice age. Changes in climate and habitat (and possibly other factors such as human hunters and major viral illnesses) led to the extinction of the mega-fauna. When prey species goes extinct, specialised predators such as sabre-tooth cats also become extinct. Those that could adapt to hunting smaller, more agile prey (e.g. hoofed grazing animals) went on to become modern cats. The population levels of elephants, our largest land mammals, are probably too small to sustain a population of sabre-toothed predators. There is no need for modern big cats to evolve into sabre-tooth forms; it would expend energy on growing the huge teeth but gain no competitive edge over other predators.
The modern Clouded Leopard is the closest we have to a sabre-tooth cat. It has the longest canines proportional to body size of any of the modern cats - the length of the fangs in approximately three times greater than the width of the fang at the socket. The backs of the canine teeth are very sharp, like those of the prehistoric sabre-toothed cats. There is a large gap between the canines and premolars, enabling them to take large bits out of their prey. Given a few million years (if humans don't wipe it out) in which to evolve and the right selective pressures, Genus Neofelis could give rise to the next generation of sabre-tooths.
There are cryptozoological reports of sabre-tooth cats surviving to the modern day in remote areas. The Tigre de Montage (Mountain Tiger) of northern Chad is described as lion-sized, striped reddish and white, tailless (or bobtailed) and having huge projecting fangs. From a selection of images, the one chosen by a Zagaoua hunter was Machairodus, an African sabre-tooth officially extinct for the last million years. The region is remote, mountainous and not well-known in zoological terms. Similar tales have come from the mountainous regions of Ecuador, Columbia, and Paraguay in South America, a region that has harboured marsupial sabre-tooths and eutherian sabre-tooth cats. In 1975, a big cat with 12 inch fangs was apparently killed in Paraguay; it was officially identified as a mutant Jaguar and unofficially identified as a Smilodon (the carcass seems to have been lost, preventing further study). Without firm evidence, e.g. a fresh or living specimen, sabre-tooth cats remain officially extinct.
Nimravids
The saber-toothed cat is one of the most famous of prehistoric icons, but perhaps one of the most neglected when it comes to public understanding. While we know dinosaurs by their genus names (names like Tyrannosaurus, Apatosaurus, and Ankylosaurus are easily come to mind), few people are familiar with saber-toothed cat genera like Smilodon, Metailurus, Dinofelis, or Xenosmilus (and there are many more). What’s even more confusing is that what we often call a saber-toothed cat is not really a cat at all, but a related carnivore called a nimravid that was molded by a striking trend in parallel evolution.
Up until a few months ago, I have never even heard of the term "nimravid", and I was quite surprised to find out that Barbourofelis and Hoplophoneus, two creatures I had always assumed were just another kind of saber-toothed cat, could not be called true cats at all. Skulls of these two genera (or manufactured facsimiles) usually sit in the same displays as those of Smilodon and other more-familiar saber-tooths, and I never thought twice to look for differences. How careless I was not to pay attention, and how careless of museums to keep lumping the remains of these separate lineages together with minimal comment.
Part of the problem with tracing the evolutionary history of mammalian carnivores is that they have generated an amazing amount of different forms; there is much diversity and plenty of branches, so every new fossil certainly can shake the tree. To keep things simple, however, all living carnivores evolved from a line of primitive carnivorous mammals called Miacids, with the Order Carnivora first becoming recognizable sometime during the Eocene (approx. 56-34 million years ago), the groups giving rise to modern dogs (Family Canidae) and cats (Family Felidae) diverging about 43 million years ago. Not all the groups that arose from the first true carnivores left living descendants, however, and such is the case with the nimravids.
So, what makes a nimravid a nimravid? They look awfully like cats, so why aren’t they lumped into the Family Felidae? What makes such distinctions so difficult is that those looking upon the skull of Smilodon and Eusmilus would have to be relatively well-versed in scientific jargon and anatomy in order to point out the most important differences. While some nimravids (like Eusmilus) had large canines, their teeth alone are not diagnostic, and the original factors used by E.D. Cope that differentiated these animals from "true" cats were the "alisphenoid canal, postglenoid foramen, carotid, posterior lacerate, and condyloid foramina, postparietal foramina" in the skull (Hunt, 1987). The various canals and foramina listed dictate the paths of various nerves and blood vessels in the skull, and the arrangement in nimravid skulls seem to be more primitive compared with true felids. Likewise, nimravids lack a two-chambered auditory bulla, which is a rounded bit of bone associated with the ear which true cats have (here’s a diagram of a dog skull pointing out the location of the bulla).
There are a few more obvious giveaways when dealing with some nimravids, however. Nimravids equipped with long canines often have more cone-shaped canines than saber-toothed cat canines (which are flatter in cross-section), and many have bony "sheaths" extending from the lower jaw into which the massive teeth fit. Perhaps the most famous example of this kind of arrangement is the genus Barbourofelis, an animal that has actually been assigned to its own family as it is likely more closely related to true cats than nimravids (Barbourofelis was previously classified as a nimravid). Because of this (and the fact that another cat-like offshoot, the marsupial Thylacosmilus) the tooth-sheath shouldn’t be considered diagnostic of nimravids only, but it does give you a substantial clue that you’re probably not dealing with an actual saber-toothed felid.
Skulls (mandibles not pictured) of 4 "saber-toothed" mammals from "The Function of Saber-Like Canines in Carnivorous Mammals" by G.G. Simpson, American Museum Novitiates, August 4, 1941. Pictured are A) Machairodus (felid), B) Hoplophoneus (nimravid), C) Smilodon (felid), and D) Thylacosmilus (marsupial).
You can see how complicated things can get; three different groups of animals arriving on the same body form from the same group of ancestors within a short amount of time. Indeed, saber-teeth seem to be a very common consequence for carnivores in this particular group, and oddly enough some living herbivores like the Musk Deer have impressive fangs as well. I’m not well-versed in evo-devo, but perhaps studying why musk deer develop such impressive teeth might give us some clues as to how it happened in these extinct cats, despite different ancestry. I should also perhaps mention that I’m curious about any sexual dimorphism between male and female saber-bearers; could sexual selection had a role in the extension of these massive canines? I don’t think it’s unreasonable to think so, especially if (as we’ll discuss) they were so long that they seemed to make these carnivores even more specialized in hunting, feeding, and social behavior than living carnivores.
Given the prevalence of massive canines amongst extinct felids and other groups, it’s a wonder why there are none living today (it should be noted, however, the Clouded Leopards have very long and impressive canines, even though they don’t peek out of their mouths when closed). It should also be noted that I have essentially left out a number of other, more distantly related saber-toothed carnivorous mammals called creodonts, which held saber-toothed hyenas like Hyaenodon in its ranks. For a time, it must have seemed like everyone and their mother had impressive fangs, and I can only wonder as to how these impressive structures became so-widespread.
It is not enough to merely say that nimravids are different, however; if they are not true saber-toothed felids, how closely are the groups related? Initially, some scientists thought that nimravids were ancestral to true cats based upon their more-primitive skull structure. As more fossils came out of the ground, the hypothesis that nimravids are closely related to true cats without being ancestral to them became favored, but this was overturned by the idea that nimravids and true cats are not very closely related, the nimravids diverging from the line that led to cats much earlier. This third view seems to make the most sense given the current fossil evidence, but I have to wonder how the reassignment of Barbourofelis will affect things, especially if it’s considered to be closer to felids than nimravids.
I included the "ancestral line" label in order to enforce the changing ideas about how evolution works, as well. In the first example the animals just kept evolving in the same line (they were the same genetic line, just with different species names as we came across them in the fossil record), but the third diagram shows that just because a new branch emerges does not mean that the ancestral line stops immediately. I have omitted Thylacosmilus and Barbourofelis as to keep things as simple as possible, and the fact that whatever I came up with would merely be a guess. I would also be remiss if I did not point this fact; while true saber-toothed cats do belong to the Family Felidae, they are all grouped together in the Subfamily Machairodontinae and do not have any living descendants. They diverged fairly early during felid evolution, ultimately becoming extinct, and I have hence tried to avoid the term "saber-toothed tiger" as much as possible. Because I’m trying to focus on nimravids for this entry I will keep the designation of "felids" for true saber-toothed cats, and hopefully I’ll eventually write a piece with more detail about the more well-known carnivores.
The big question involving these animals, however, is "How in the hell did they actually use those teeth?" Given that saber-toothed mammalian predators evolved three times in a geologically short time in three separate groups of predators suggests that they were useful for something, but how do you bite with teeth that extend past your lower jaw? In considering this question, it’s important to remember that when biting only the lower jaw is actually moving, so if a saber-toothed mammal wanted to impale a prey item with its long canines, it would have to throw its neck around with considerable force to achieve that end. In fact, this kind of action has already been proposed by some, the dynamics of felid saber-tooth skulls making it difficult to conceive how such huge canines could be used to effectively bite prey.
Part of the problem with having saber-teeth is that you need to open your jaw exceedingly wide in order to get food in your mouth. The oft-cited measurement for the gape of the felid Smilodon is 120 degrees (no source I’ve seen references where this measurement came from), and even if this is wrong we know that in order to get food into their mouths, many of the hyper-saber-toothed mammals would need to open their jaws to a 90 degree angle or more, otherwise they would not be able to get food in their mouths. What this means, as far as muscle strength is concerned, is that the muscles would not be as strong as in other cats, getting the mouth open being more important to a strong bite, so saber-toothed mammals would not have the crushing power of modern tigers or lions. Likewise, owning saber-teeth can make hunting difficult; if you stick your teeth into a live animal and it struggles, you could very well lose a tooth. Likewise the teeth would be more fragile, so putting extreme stresses on them (like crushing bone) would largely be out of the question too; it would be more effective and safe to attack soft parts of an animal than to try for the take-down neck-bites that modern cats employ.
We should be careful in our assumptions, however; we’re dealing with extinct animals, and their method of capturing/subduing prey may have differed significantly from any living carnivore. While I just mentioned that saber-toothed mammals likely had weak jaws, a 2005 study suggests that they had jaws as strong or stronger than living big cats, with different killing strategies depending on the overall durability/robustness of the saber-teeth. Likewise, an earlier study (1996) based upon tooth wear in Smilodon was unable to match wear indicative of bone crushing/chewing/abrasion with living hyenas, canids, and cats, suggesting that Smilodon may have avoided contact with bone as much as possible. Indeed, even though all these animals had impressive canines, not all their canines were equal, and some would be better suited to dealing with stresses involved with prey capture than others. Still, I would regard many of these teeth as delicate, and I can only imagine the pain these mammals must have endured when one of them broke.
Other hypotheses about how these animals employed their teeth involves the white shark-like tactic of disemboweling the softer underbelly of prey, then waiting for the eviscerated creature to die. This would be a rather risky move, the predator essentially sticking its head right between both sets of sharp hooves (assuming the prey was an ungulate). What seems more reasonable would be a strategy based upon cooperation, much like modern lions taking down huge water buffalo. If the group could bring down the prey with their claws, one animal could deliver the killing bite to the neck, minimizing the amount of potential harm to itself. This hypothesis, however, requires the study of behavior that we are no longer privy to, and it would be unreasonable to infer such a pattern on all saber-toothed mammals as the rule.
In his own paper studying the various methods of attack saber-toothed mammals could have used, G.G. Simpson concluded that they were best adapted for stabbing, not as much for slicing (although he conceded that they likely did this as well), the dentition of these animals showing their predatory habits (it had been hypothesized earlier that these animals may have been scavengers). Simpson’s study is interesting, but prey is generally not taken into account; only the effectiveness of different strategies for ripping up the assumed prey. While it certainly serves as a good reference point from a mechanical point of view, the skulls of the animals are considered out of context, and so the major mysteries of these animals remain unsolved.
Ultimately, all the known saber-toothed predators died out, regardless of their affinities. One of the most popular views (which I am surprised to still hear) is that the teeth of these animals simply became so huge that they could not properly open and close their mouths, driving the species to extinction. If there are urban legends in paleontology, surely this is one of the most annoying and persistent. G.G. Simpson refutes this idea in his popular work The Meaning of Evolution, published more than 30 years before I had heard it from various documentaries claiming scientific accuracy;
The sabertooth is one of the most famous of animals just because it is often innocently supposed to be an indisputable example of an inadaptive trend. In fields far remote from paleontology the poor sabertooth has some to figure as a horrible example, a pathetic case history of evolution gone wrong. Its supposed evidence is thus characteristically summarized in a book on (human) personality: "The long canine tooth of the saber-toothed tiger grew more and more into an impossible occlusion. Finally, it was so long that the tiger could not bite effectively, and the animal became extinct." Now, like so many things that everyone seems to know, this is not true… Throughout their history the size of sabertooth canines varied considerably from one group to another but varied about a fairly constant average size, which is exactly what would be expected if the size were adaptive at all times and there were no secular trend in adaptive advantage but only local and temporary differences in its details. The biting mechanism in the last sabertooths was still perfectly effective, no less and probably no more so than in the Oligocene. To characterize a finally ineffective a mechanism that persisted without essential change in a group abundant and obviously highly successful for some 40,000,000 years seems quaintly illogical! In short, the "inadaptive trend" of the sabertooth is a mere fairy tale, or more fairly, it was an error based on too facile conclusion from imperfect information and it has since been perpetuated as a scientific legend.
Why saber-teeth seemed to be so trendy among predatory mammals, only to disappear entirely, I have no idea. Obviously they must have been good for something, some common developmental, ecological, or other trend driving canines to be longer, only to (perhaps) cause the animals to be so specialized that they could no longer compete with other carnivores who did not have to be so concerned about their teeth. At the very least, however, I hope this post have served to bring to attention a group generally overlooked, often mistaken for their cousins, when they have a rich evolutionary history of their own.
Sometimes when it looks like a cat and acts like a cat it’s not; it’s a nimravid! Nimravids are a rather problematic group in the order Carnivora of uncertain descent. Entering the fossil record in the late Eocene (36 million years ago), their exact relationship to other carnivore families is unresolved, but they paralleled cats to an amazing degree. With short faces and fully hooded, retractile claws, they came in body shapes and sizes that covered all of the diversity that true cats would later manifest. Some were as small as house cats, while others were the ecological and functional equivalents of lynx, cheetah and leopard. And all except one lone genus were saber-toothed.
Before we go on, let’s get these curved teeth straight. Saber-teeth evolved from normal, conical canine teeth into two forms. Dirk teeth were the long, curved, finely serrated blades that we normally think of when we think saber-toothed. The famous Smilodon - the saber-toothed "tiger" of LaBrea Tar Pits fame - was a dirk-toothed true felid. There were also scimitar-toothed animals with shorter, more coarsely serrated blades that usually evolved in conjunction with longer-legged cursorial body types. Most of the dirk-toothed animals were compact, robust stalkers.
Barbourofelis fricki was the last of the nimravids, dirk-toothed and huge. Its name, "Barbour’s Cat", shows just how close and how confusing convergent similarities can be. This was a massive creature, the size of a modern lion, but more heavily built. At rest or when walking, it stood and moved in a plantigrade fashion, rather like a bear. One look at the business end of the animal however, lets you know that Barbourofelis wasn’t looking for berries. It possessed some of the largest saber-teeth of all time, and the body to back them up. Behind the catlike face, the head had a flat profile caused by the expanded occipital region at the back of the skull. This allowed for the attachment of powerful neck and shoulder musculature, the driving force behind the sabers. This was a stalker and killer of large game.
Nimravids have an interesting history in North America. Throughout the Oligocene, they assumed all the predatory roles later taken by true cats. But by the beginning of the Miocene, 23 million years ago, they had all disappeared. For whatever reason, perhaps because there was an abundance of arctoid (doglike) carnivores of all descriptions present here at the time, there were no nimravids in North America for the next few million years. Felids also were missing and made their North American debut in the form of a small conical-toothed animal, Pseudailurus, about 17 million years ago. (Showing up to really confuse things a few million years later was a scimitar-toothed true cat named Nimravides! Got that? "Barbour’s Cat" is a nimravid, and Nimravides is a real cat! Aurgh!!!)
Finally, after a 10 million year absence, nimravids returned, most likely from Asia, in the form of Barbourofelis. They show up as moderate-sized creatures with moderate-sized dirk teeth.
This is an early barbourofelid species based on California material attacking the three-toed horse Hipparion. It is also one of my very first digital paintings. It was done a few years ago for an article by Joe Eaton in the September, 2001 issue of Bay Nature, a Northern California publication dedicated to all aspects of San Francisco Bay Area natural history. Getting this image ready for Olduvai - and I did work on it - I was amazed at how much I’ve improved with this computerized medium. The drawing is fine, except that I gave Barbourofelis a tail that was too long (digitally fixed now), but the color rendering was primitive. My thanks to the editor, David Loeb, for letting me experiment on his dime.
One of the problems with having killing teeth like this was eating the meat once it was secured. Barbourofelis solved the problem by having robust incisors and shearing carnassial teeth to strip and slice meat from bone. Utilizing the carnassials must have been problematic, because the animal had to move its cheek out of the way behind and outside the sabers. You don’t have a 5 million year history without solving those gnawing problems. (I’m SO sorry! I couldn’t stop myself.)
The trouble with a post like this is that it’s a book in the making. There is so much to draw and write about, and so many questions yet to be answered, that I really don’t know what to leave out and what tidbit people might find most interesting. For instance, looking at the paintings the one thing you can’t help but notice is the dependent flange on the mandible of the animals. These protected the sabers when the mouth was closed and there was little room to spare between the saber and the bone of the lower jaw. Maybe I’ve missed it, but I can’t recall having seen anybody paint this as the gum-lined half sheath I’ve portrayed here. There certainly wasn’t enough room for hair to grow, and unless the tooth was supported by being in contact with the flange, it would have defeated its supposed purpose. (I should add that the dirk-toothed felids like Smilodon didn’t have the flange. This is such fun.)
I want to note that things will be changing here at Olduvai George soon. I’ll have an announcement next week about some exciting new developments that will help me to post more often and include more new illustrations. New old whales are coming.
I was hoping to have Kutchicetus up this week but I haven’t nailed the look yet as the new work is based on material I hadn’t seen until last week. 2007 promises to be a very busy year.
Up until a few months ago, I have never even heard of the term "nimravid", and I was quite surprised to find out that Barbourofelis and Hoplophoneus, two creatures I had always assumed were just another kind of saber-toothed cat, could not be called true cats at all. Skulls of these two genera (or manufactured facsimiles) usually sit in the same displays as those of Smilodon and other more-familiar saber-tooths, and I never thought twice to look for differences. How careless I was not to pay attention, and how careless of museums to keep lumping the remains of these separate lineages together with minimal comment.
Part of the problem with tracing the evolutionary history of mammalian carnivores is that they have generated an amazing amount of different forms; there is much diversity and plenty of branches, so every new fossil certainly can shake the tree. To keep things simple, however, all living carnivores evolved from a line of primitive carnivorous mammals called Miacids, with the Order Carnivora first becoming recognizable sometime during the Eocene (approx. 56-34 million years ago), the groups giving rise to modern dogs (Family Canidae) and cats (Family Felidae) diverging about 43 million years ago. Not all the groups that arose from the first true carnivores left living descendants, however, and such is the case with the nimravids.
So, what makes a nimravid a nimravid? They look awfully like cats, so why aren’t they lumped into the Family Felidae? What makes such distinctions so difficult is that those looking upon the skull of Smilodon and Eusmilus would have to be relatively well-versed in scientific jargon and anatomy in order to point out the most important differences. While some nimravids (like Eusmilus) had large canines, their teeth alone are not diagnostic, and the original factors used by E.D. Cope that differentiated these animals from "true" cats were the "alisphenoid canal, postglenoid foramen, carotid, posterior lacerate, and condyloid foramina, postparietal foramina" in the skull (Hunt, 1987). The various canals and foramina listed dictate the paths of various nerves and blood vessels in the skull, and the arrangement in nimravid skulls seem to be more primitive compared with true felids. Likewise, nimravids lack a two-chambered auditory bulla, which is a rounded bit of bone associated with the ear which true cats have (here’s a diagram of a dog skull pointing out the location of the bulla).
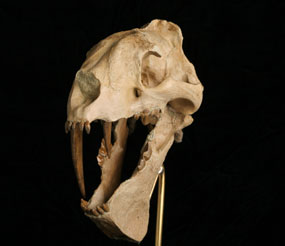{kind=link}
{kind=link}
There are a few more obvious giveaways when dealing with some nimravids, however. Nimravids equipped with long canines often have more cone-shaped canines than saber-toothed cat canines (which are flatter in cross-section), and many have bony "sheaths" extending from the lower jaw into which the massive teeth fit. Perhaps the most famous example of this kind of arrangement is the genus Barbourofelis, an animal that has actually been assigned to its own family as it is likely more closely related to true cats than nimravids (Barbourofelis was previously classified as a nimravid). Because of this (and the fact that another cat-like offshoot, the marsupial Thylacosmilus) the tooth-sheath shouldn’t be considered diagnostic of nimravids only, but it does give you a substantial clue that you’re probably not dealing with an actual saber-toothed felid.
{kind=link}
{kind=link}
Skulls (mandibles not pictured) of 4 "saber-toothed" mammals from "The Function of Saber-Like Canines in Carnivorous Mammals" by G.G. Simpson, American Museum Novitiates, August 4, 1941. Pictured are A) Machairodus (felid), B) Hoplophoneus (nimravid), C) Smilodon (felid), and D) Thylacosmilus (marsupial).
You can see how complicated things can get; three different groups of animals arriving on the same body form from the same group of ancestors within a short amount of time. Indeed, saber-teeth seem to be a very common consequence for carnivores in this particular group, and oddly enough some living herbivores like the Musk Deer have impressive fangs as well. I’m not well-versed in evo-devo, but perhaps studying why musk deer develop such impressive teeth might give us some clues as to how it happened in these extinct cats, despite different ancestry. I should also perhaps mention that I’m curious about any sexual dimorphism between male and female saber-bearers; could sexual selection had a role in the extension of these massive canines? I don’t think it’s unreasonable to think so, especially if (as we’ll discuss) they were so long that they seemed to make these carnivores even more specialized in hunting, feeding, and social behavior than living carnivores.
Given the prevalence of massive canines amongst extinct felids and other groups, it’s a wonder why there are none living today (it should be noted, however, the Clouded Leopards have very long and impressive canines, even though they don’t peek out of their mouths when closed). It should also be noted that I have essentially left out a number of other, more distantly related saber-toothed carnivorous mammals called creodonts, which held saber-toothed hyenas like Hyaenodon in its ranks. For a time, it must have seemed like everyone and their mother had impressive fangs, and I can only wonder as to how these impressive structures became so-widespread.
It is not enough to merely say that nimravids are different, however; if they are not true saber-toothed felids, how closely are the groups related? Initially, some scientists thought that nimravids were ancestral to true cats based upon their more-primitive skull structure. As more fossils came out of the ground, the hypothesis that nimravids are closely related to true cats without being ancestral to them became favored, but this was overturned by the idea that nimravids and true cats are not very closely related, the nimravids diverging from the line that led to cats much earlier. This third view seems to make the most sense given the current fossil evidence, but I have to wonder how the reassignment of Barbourofelis will affect things, especially if it’s considered to be closer to felids than nimravids.
I included the "ancestral line" label in order to enforce the changing ideas about how evolution works, as well. In the first example the animals just kept evolving in the same line (they were the same genetic line, just with different species names as we came across them in the fossil record), but the third diagram shows that just because a new branch emerges does not mean that the ancestral line stops immediately. I have omitted Thylacosmilus and Barbourofelis as to keep things as simple as possible, and the fact that whatever I came up with would merely be a guess. I would also be remiss if I did not point this fact; while true saber-toothed cats do belong to the Family Felidae, they are all grouped together in the Subfamily Machairodontinae and do not have any living descendants. They diverged fairly early during felid evolution, ultimately becoming extinct, and I have hence tried to avoid the term "saber-toothed tiger" as much as possible. Because I’m trying to focus on nimravids for this entry I will keep the designation of "felids" for true saber-toothed cats, and hopefully I’ll eventually write a piece with more detail about the more well-known carnivores.
The big question involving these animals, however, is "How in the hell did they actually use those teeth?" Given that saber-toothed mammalian predators evolved three times in a geologically short time in three separate groups of predators suggests that they were useful for something, but how do you bite with teeth that extend past your lower jaw? In considering this question, it’s important to remember that when biting only the lower jaw is actually moving, so if a saber-toothed mammal wanted to impale a prey item with its long canines, it would have to throw its neck around with considerable force to achieve that end. In fact, this kind of action has already been proposed by some, the dynamics of felid saber-tooth skulls making it difficult to conceive how such huge canines could be used to effectively bite prey.
Part of the problem with having saber-teeth is that you need to open your jaw exceedingly wide in order to get food in your mouth. The oft-cited measurement for the gape of the felid Smilodon is 120 degrees (no source I’ve seen references where this measurement came from), and even if this is wrong we know that in order to get food into their mouths, many of the hyper-saber-toothed mammals would need to open their jaws to a 90 degree angle or more, otherwise they would not be able to get food in their mouths. What this means, as far as muscle strength is concerned, is that the muscles would not be as strong as in other cats, getting the mouth open being more important to a strong bite, so saber-toothed mammals would not have the crushing power of modern tigers or lions. Likewise, owning saber-teeth can make hunting difficult; if you stick your teeth into a live animal and it struggles, you could very well lose a tooth. Likewise the teeth would be more fragile, so putting extreme stresses on them (like crushing bone) would largely be out of the question too; it would be more effective and safe to attack soft parts of an animal than to try for the take-down neck-bites that modern cats employ.
We should be careful in our assumptions, however; we’re dealing with extinct animals, and their method of capturing/subduing prey may have differed significantly from any living carnivore. While I just mentioned that saber-toothed mammals likely had weak jaws, a 2005 study suggests that they had jaws as strong or stronger than living big cats, with different killing strategies depending on the overall durability/robustness of the saber-teeth. Likewise, an earlier study (1996) based upon tooth wear in Smilodon was unable to match wear indicative of bone crushing/chewing/abrasion with living hyenas, canids, and cats, suggesting that Smilodon may have avoided contact with bone as much as possible. Indeed, even though all these animals had impressive canines, not all their canines were equal, and some would be better suited to dealing with stresses involved with prey capture than others. Still, I would regard many of these teeth as delicate, and I can only imagine the pain these mammals must have endured when one of them broke.
Other hypotheses about how these animals employed their teeth involves the white shark-like tactic of disemboweling the softer underbelly of prey, then waiting for the eviscerated creature to die. This would be a rather risky move, the predator essentially sticking its head right between both sets of sharp hooves (assuming the prey was an ungulate). What seems more reasonable would be a strategy based upon cooperation, much like modern lions taking down huge water buffalo. If the group could bring down the prey with their claws, one animal could deliver the killing bite to the neck, minimizing the amount of potential harm to itself. This hypothesis, however, requires the study of behavior that we are no longer privy to, and it would be unreasonable to infer such a pattern on all saber-toothed mammals as the rule.
In his own paper studying the various methods of attack saber-toothed mammals could have used, G.G. Simpson concluded that they were best adapted for stabbing, not as much for slicing (although he conceded that they likely did this as well), the dentition of these animals showing their predatory habits (it had been hypothesized earlier that these animals may have been scavengers). Simpson’s study is interesting, but prey is generally not taken into account; only the effectiveness of different strategies for ripping up the assumed prey. While it certainly serves as a good reference point from a mechanical point of view, the skulls of the animals are considered out of context, and so the major mysteries of these animals remain unsolved.
Ultimately, all the known saber-toothed predators died out, regardless of their affinities. One of the most popular views (which I am surprised to still hear) is that the teeth of these animals simply became so huge that they could not properly open and close their mouths, driving the species to extinction. If there are urban legends in paleontology, surely this is one of the most annoying and persistent. G.G. Simpson refutes this idea in his popular work The Meaning of Evolution, published more than 30 years before I had heard it from various documentaries claiming scientific accuracy;
The sabertooth is one of the most famous of animals just because it is often innocently supposed to be an indisputable example of an inadaptive trend. In fields far remote from paleontology the poor sabertooth has some to figure as a horrible example, a pathetic case history of evolution gone wrong. Its supposed evidence is thus characteristically summarized in a book on (human) personality: "The long canine tooth of the saber-toothed tiger grew more and more into an impossible occlusion. Finally, it was so long that the tiger could not bite effectively, and the animal became extinct." Now, like so many things that everyone seems to know, this is not true… Throughout their history the size of sabertooth canines varied considerably from one group to another but varied about a fairly constant average size, which is exactly what would be expected if the size were adaptive at all times and there were no secular trend in adaptive advantage but only local and temporary differences in its details. The biting mechanism in the last sabertooths was still perfectly effective, no less and probably no more so than in the Oligocene. To characterize a finally ineffective a mechanism that persisted without essential change in a group abundant and obviously highly successful for some 40,000,000 years seems quaintly illogical! In short, the "inadaptive trend" of the sabertooth is a mere fairy tale, or more fairly, it was an error based on too facile conclusion from imperfect information and it has since been perpetuated as a scientific legend.
Why saber-teeth seemed to be so trendy among predatory mammals, only to disappear entirely, I have no idea. Obviously they must have been good for something, some common developmental, ecological, or other trend driving canines to be longer, only to (perhaps) cause the animals to be so specialized that they could no longer compete with other carnivores who did not have to be so concerned about their teeth. At the very least, however, I hope this post have served to bring to attention a group generally overlooked, often mistaken for their cousins, when they have a rich evolutionary history of their own.
Sometimes when it looks like a cat and acts like a cat it’s not; it’s a nimravid! Nimravids are a rather problematic group in the order Carnivora of uncertain descent. Entering the fossil record in the late Eocene (36 million years ago), their exact relationship to other carnivore families is unresolved, but they paralleled cats to an amazing degree. With short faces and fully hooded, retractile claws, they came in body shapes and sizes that covered all of the diversity that true cats would later manifest. Some were as small as house cats, while others were the ecological and functional equivalents of lynx, cheetah and leopard. And all except one lone genus were saber-toothed.
Before we go on, let’s get these curved teeth straight. Saber-teeth evolved from normal, conical canine teeth into two forms. Dirk teeth were the long, curved, finely serrated blades that we normally think of when we think saber-toothed. The famous Smilodon - the saber-toothed "tiger" of LaBrea Tar Pits fame - was a dirk-toothed true felid. There were also scimitar-toothed animals with shorter, more coarsely serrated blades that usually evolved in conjunction with longer-legged cursorial body types. Most of the dirk-toothed animals were compact, robust stalkers.
Barbourofelis fricki was the last of the nimravids, dirk-toothed and huge. Its name, "Barbour’s Cat", shows just how close and how confusing convergent similarities can be. This was a massive creature, the size of a modern lion, but more heavily built. At rest or when walking, it stood and moved in a plantigrade fashion, rather like a bear. One look at the business end of the animal however, lets you know that Barbourofelis wasn’t looking for berries. It possessed some of the largest saber-teeth of all time, and the body to back them up. Behind the catlike face, the head had a flat profile caused by the expanded occipital region at the back of the skull. This allowed for the attachment of powerful neck and shoulder musculature, the driving force behind the sabers. This was a stalker and killer of large game.
Nimravids have an interesting history in North America. Throughout the Oligocene, they assumed all the predatory roles later taken by true cats. But by the beginning of the Miocene, 23 million years ago, they had all disappeared. For whatever reason, perhaps because there was an abundance of arctoid (doglike) carnivores of all descriptions present here at the time, there were no nimravids in North America for the next few million years. Felids also were missing and made their North American debut in the form of a small conical-toothed animal, Pseudailurus, about 17 million years ago. (Showing up to really confuse things a few million years later was a scimitar-toothed true cat named Nimravides! Got that? "Barbour’s Cat" is a nimravid, and Nimravides is a real cat! Aurgh!!!)
Finally, after a 10 million year absence, nimravids returned, most likely from Asia, in the form of Barbourofelis. They show up as moderate-sized creatures with moderate-sized dirk teeth.
This is an early barbourofelid species based on California material attacking the three-toed horse Hipparion. It is also one of my very first digital paintings. It was done a few years ago for an article by Joe Eaton in the September, 2001 issue of Bay Nature, a Northern California publication dedicated to all aspects of San Francisco Bay Area natural history. Getting this image ready for Olduvai - and I did work on it - I was amazed at how much I’ve improved with this computerized medium. The drawing is fine, except that I gave Barbourofelis a tail that was too long (digitally fixed now), but the color rendering was primitive. My thanks to the editor, David Loeb, for letting me experiment on his dime.
One of the problems with having killing teeth like this was eating the meat once it was secured. Barbourofelis solved the problem by having robust incisors and shearing carnassial teeth to strip and slice meat from bone. Utilizing the carnassials must have been problematic, because the animal had to move its cheek out of the way behind and outside the sabers. You don’t have a 5 million year history without solving those gnawing problems. (I’m SO sorry! I couldn’t stop myself.)
The trouble with a post like this is that it’s a book in the making. There is so much to draw and write about, and so many questions yet to be answered, that I really don’t know what to leave out and what tidbit people might find most interesting. For instance, looking at the paintings the one thing you can’t help but notice is the dependent flange on the mandible of the animals. These protected the sabers when the mouth was closed and there was little room to spare between the saber and the bone of the lower jaw. Maybe I’ve missed it, but I can’t recall having seen anybody paint this as the gum-lined half sheath I’ve portrayed here. There certainly wasn’t enough room for hair to grow, and unless the tooth was supported by being in contact with the flange, it would have defeated its supposed purpose. (I should add that the dirk-toothed felids like Smilodon didn’t have the flange. This is such fun.)
I want to note that things will be changing here at Olduvai George soon. I’ll have an announcement next week about some exciting new developments that will help me to post more often and include more new illustrations. New old whales are coming.
I was hoping to have Kutchicetus up this week but I haven’t nailed the look yet as the new work is based on material I hadn’t seen until last week. 2007 promises to be a very busy year.